Индукция ферментов
Как было показано выше, с возрастом меняется не только активность ферментов - в определенные периоды жизни меняется также набор изоферментов. Отсюда возникают следующие вопросы:
1) Являются ли эти изменения необратимыми?
2) Возможно ли эти изменения направить вспять и можем ли мы регулировать их уровни в различные периоды жизни?
3) Какие факторы ответственны за эти изменения?
4) Какое значение эти изменения имеют для различных функций организма и для старения?
Получен ряд данных, позволяющих ответить на часть этих вопросов. Известно, что вторичные клетки флоэмы моркови, культивируемые in vitro, дают начало целому растению [107]. Следовательно, высокодифференцированные клетки флоэмы подвергаются дополнительной дифференциации, давая начало клеткам новых типов. На животных клетках подобные эксперименты пока не проводились. Митохондриальная малатдегидрогеназа (мМДГ) индуцируется кортизоном в печени молодых крыс, у которых удален надпочечник, и не индуцируется у старых [53]. Однако, если печень старой крысы регенерирует в результате гепатэктомии, то кортизон вызывает индукцию мМДГ. Таким образом, в этом случае способность к индукции восстанавливается и печень старой крысы функционирует так же, как печень молодой. Из данных экспериментов следует, что изменения макромолекул, происходящие при старении, не являются необратимыми. Содержание ферментов, понизившееся в старости, может быть увеличено путем индукции, и повреждения таким образом могут быть устранены.
Известно, что индукция, или повышение содержания ферментов, вызывается несколькими индукторами, или эффекторами. Индукция является специфическим ответом на специфический стимул, или индуктор, которым может быть субстрат, гормон, метаболит или даже экзогенный фактор - например температура, т. е. индукция - это адаптивный процесс. Как известно, адаптивность организма к условиям окружающей среды в старости снижается. Поэтому представляется интересным исследовать изменения адаптивности или индуцибельности ферментов с возрастом. Поскольку каждый фермент кодируется специфическим геном, подобные исследования могут помочь в выяснении таких функций специфических генов, как транскрипция специфических мРНК и регуляция этой транскрипции. Проводя их, можно также решить вопрос о том, происходят или не происходят изменения в экспрессии генов в старости.
В работах по индукции синтеза ферментов в тканях крысы путем введения различных гормонов было показано, что при одинаковых условиях эксперимента индукция у старых крыс в общем ниже, чем у животных в зрелом возрасте. Такие результаты были получены для глюкозо-6-фосфатазы и фруктозобисфосфат-альдолазы [101], фосфофруктокиназы [102, 103] и гексозофосфат-изомеразы [103], пируваткиназы [21], малатдегидрогеназы [53] и ацетилхолинэстеразы [81]. Для нескольких ферментов в зрелом возрасте и в старости получены одинаковые значения индукции - это триптофанпирролаза [42], аланинаминотрансфераза (ААТ) [87] и тирозинаминотрансфераза (ТАТ) [91], а индукция аргиназы [90] у старых животных оказалась выше (табл. 3.4), чем у животных в среднем возрасте.
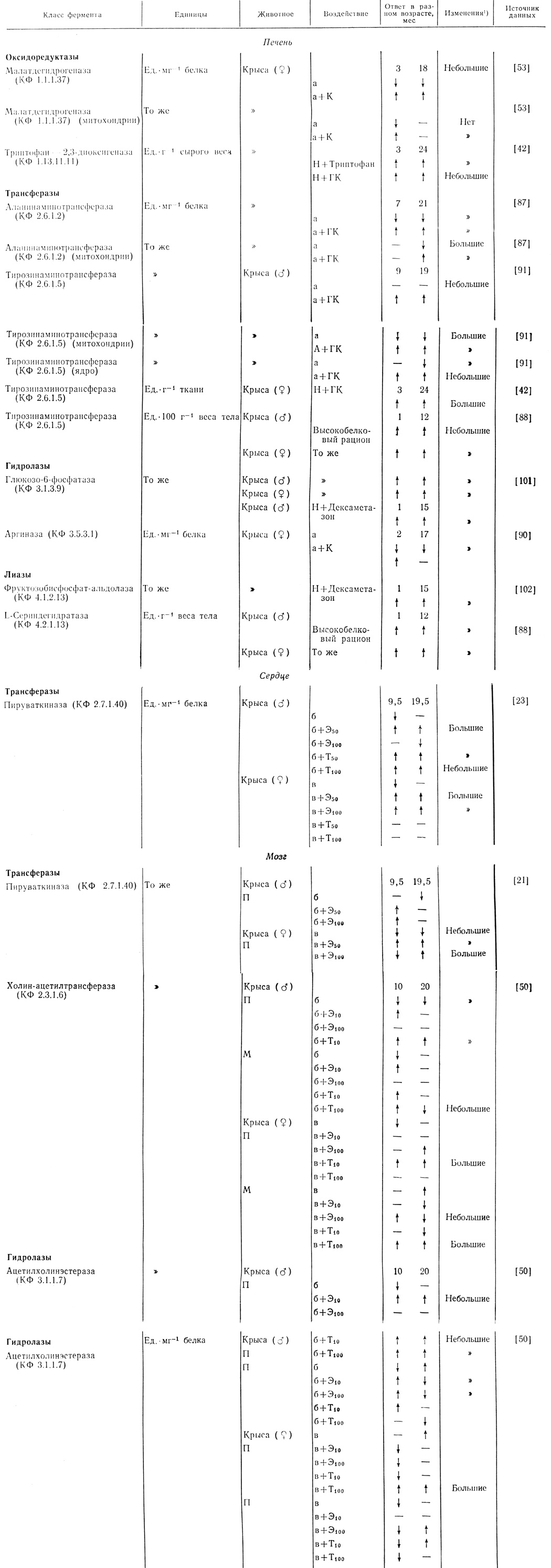
Таблица 3.4. Индукция ферментов
Обозначения в Таблице: ↓ уменьшение; ↑ увеличение; "-" нет эффекта; Н - норма; К - кортизон; ГК - гидрокортизон; а - удаление надпочечника; б - удаление семенников; в - удаление яичников; Э50-Э100 - введение эстрадиола в количестве 50 и 100 мг·г-1 веса тела; Т50 и Т100 - введение тестостерона в количестве 50 и 100 мг·г-1 веса тела; П - полушария головного мозга; М - мозжечок.
Некоторые ферменты, например глюкокиназа и тирозинаминотрансфераза из печени мыши, индуцируются при голодании [2]. Было показано также, что тирозинаминотрансфераза печени индуцируется при низкой температуре [33]. В этих исследованиях установлено, что индукция у старых и молодых животных одинакова, но у первых время, необходимое для индукции, больше (табл. 3.4).
В общем, индукция оксидоредуктаз в старости понижается, а трансфераз - повышается или не изменяется. Индукция гидролаз и лиаз печени млекопитающих в старческом возрасте понижается, так же как и индукция трансфераз и гидролаз мозга. Однако пока число исследованных ферментов невелико, нельзя с уверенностью сделать заключение о закономерностях индукции определенных ферментов.
В приведенных исследованиях было показано также, что индукция одного и того же фермента по-разному меняется в разных органах. Например, индукция пируваткиназы в сердце старой крысы выше, чем у животного в среднем возрасте, в то время как в мозгу старой крысы она ниже по сравнению с животным в среднем возрасте [21-23].
Индукция многих ферментов в старческом возрасте уменьшается, и поэтому ослабевает адаптивность организма или его способность к ответу на соответствующий стимул. Эти ответы на основании времени, затрачиваемого на индукцию, и величины индукции (рис. 3.7) подразделяют на четыре главных типа [5]. У типа 1 лаг-период индукции в старости больше, но ее величина не меняется, если индуктор применяется в течение достаточного времени. Примерами этого типа ферментов являются глюкокиназа [1] (рис. 3.8), ДНК-полимераза [3] (рис. 3.9), тирозинаминотрансфераза [2] и сериндегидратаза [88] печени крыс при ответе на действие глюкозы, АКТГ и кортизона соответственно [5]. Лаг-период индукции тирозинаминотрансферазы после выдерживания при низкой температуре у старой мыши также увеличен [33].
![Рис. 3.7. Зависимость ферментной адаптации от возраста [15]. В общем виде показаны четыре основных типа влияния старения на время протекания и величину гипотетических адаптивных изменений активности ферментов. Сплошная линия - молодые животные, штриховая - среднего возраста, пунктирная - старые](pic/000042.jpg)
Рис. 3.7. Зависимость ферментной адаптации от возраста [15]. В общем виде показаны четыре основных типа влияния старения на время протекания и величину гипотетических адаптивных изменений активности ферментов. Сплошная линия - молодые животные, штриховая - среднего возраста, пунктирная - старые
![Рис. 3.8. Зависимость лаг-периода индукции глюкокиназы от возраста крысы [4]](pic/000043.jpg)
Рис. 3.8. Зависимость лаг-периода индукции глюкокиназы от возраста крысы [4]
![Рис. 3.9. Возрастные различия начала и максимума скорости синтеза ДНК в подчелюстной железе крысы после инъекции изопротеренола [3]. 1 - возраст 2 мес; 2 - возраст 12 мес](pic/000044.jpg)
Рис. 3.9. Возрастные различия начала и максимума скорости синтеза ДНК в подчелюстной железе крысы после инъекции изопротеренола [3]. 1 - возраст 2 мес; 2 - возраст 12 мес
У ферментов типа 2 степень индукции при одинаковой дозе индуктора в старческом возрасте или больше, или меньше. Индукция орнитин-оксокислота - аминотрансферазы кортизоном, глюкозо-6-фосфатазы в печени высокобелковым рационом [88], цитоплазматической малатдегидрогеназы адреналэктомированных крыс кортизоном [53] и орнитиндекарбоксилазы дексаметазоном [30] в старости значительно понижается. Экстремальные случаи такого понижения наблюдаются для митохондриальной малатдегидрогеназы [53] и аргиназы [90] печени после удаления надпочечника и введения кортизона и для ацетилхолинэстеразы мозга крысы [81] после овариэктомии и введения эстрадиола. Во всех этих случаях у старых животных наблюдается полное отсутствие индукции. Вместе с тем индукция глутаминсинтетазы кортизоном у старых крыс больше и в норме, и после удаления надпочечника [89]. Индукция в этих случаях исследовалась только одномоментно, и лаг-период ни для старых крыс, ни для крыс в среднем возрасте неизвестен.
У ферментов типа 3 в старости происходят изменения и величины индукции, и времени, необходимого для индукции. К этому типу относятся замедленное и более слабое повышение активности тимидинкиназы и дезокситимидилатсинтетазы из слюнных желез у крысы после введения изопротеринола [96] или у дрозофилы после обработки аминотриазолом [82].
Для ферментов типа 4 ответ на действие индуктора в среднем возрасте и в старости одинаков. К этому типу относится индукция глюкокиназы, тирозинаминотрансферазы [6] и митохондриальной глицерол-3-фосфат - дегидрогеназы [17] инсулином, кортизоном и тироксином соответственно.
Как и в случае содержания ферментов у нормальных животных, способность различных ферментов к индукции разными индукторами зависит, по-видимому, от линии и пола животных, времени измерения активности фермента, условий, в которых содержались животные, и их физиологического состояния. Однако очевидно, что в одной и той же популяции индукция некоторых ферментов может меняться. При этом может происходить изменение или величины, или лаг-периода, или того и другого вместе. Последовательность событий, начинающаяся с момента проникновения стероидного гормона в клетку-мишень и завершающаяся синтезом специфического белка, более или менее установлена. Гормон проникает в клетку-мишень и связывается со специфическим белковым рецептором в цитозоле, образуя комплекс гормон - рецептор (Г-Р). Этот комплекс проникает в ядро, где связывается со специфическими акцепторными центрами хроматина и стимулирует транскрипцию, после чего происходит синтез специфического белка (рис. 5.2). Необходимо точно определить то место (места) в этой последовательности событий, на котором в старческом возрасте происходит нарушение индукции стероидными гормонами.
Такая попытка была предпринята Канунго и его сотрудниками [54], которые показали, что эстрадиол накапливается в мозгу крысы. Введение эстрадиола (10 мкг/100 г) индуцирует ацетилхолинэстеразу в полушариях головного мозга незрелых и зрелых крыс после овариэктомии, но не влияет на синтез этого фермента у старых крыс [81]. Одной из возможных причин ухудшения индукции ацетилхолинэстеразы эстрадиолом в мозгу старой крысы может быть понижение содержания эстрадиол-связывающих рецепторных белков. Чтобы проверить это предположение, гомогенаты, полученные из полушария головного мозга 7-, 44- и 108-недельных крыс с удаленными яичниками, инкубировали с 3Н-эстрадиолом и количественно определяли эстрадиол-специфичный белок. Оказалось, что самое большое сродство этого белка к 17β-эстрадиолу характерно для головного мозга незрелых крыс, у которых наблюдается и самая сильная индукция ацетилхолинэстеразы (рис. 5.6) [56]. Сродство белка к эстрадиолу с возрастом понижается. Следовательно, одной из молекулярных причин ухудшения индукции ацетилхолинэстеразы эстрадиолом, по-видимому, является уменьшение содержания эстрадиол-специфического белка. Возникает вопрос: чем же вызывается это уменьшение? Известно, что эстрадиол стимулирует синтез своего собственного рецептора. Является ли понижение содержания специфического рецептора эстрадиола в мозгу крыс-самок следствием понижения содержания эстрадиола, которое, как известно, происходит в старости? У крыс-самцов уровень андрогена в старости также снижается. Влияет ли это на содержание ацетилхолинэстеразы в полушариях головного мозга? Если такое влияние существует, то для повышения содержания ацетилхолинэстеразы, жизненно важной для мозга, старым животным можно вводить стероидные гормоны.
Попытки повысить уровень определенных ферментов в мозгу старых крыс путем введения стероидных гормонов дали обнадеживающие результаты. Кастрация старых мужских и женских особей крыс понижает содержание в полушариях мозга двух жизненно важных ферментов, ответственных за проводимость нервов, - ацетилхолинэстеразы и холин-ацетилтрансферазы. Ацетилхолинэстераза катализирует гидролиз ацетилхолина, а холин-ацетилтрансфераза - его синтез. Введение 10 мкг тестостерона на 100 г веса повышает содержание того и другого фермента у старых крыс обоих полов приблизительно до уровня репродуктивного периода. В отношении индукции холин-ацетилтрансферазы тестостерон более эффективен, чем эстрадиол (рис. 5.9) [50]. Такие же результаты были получены для пируваткиназы мозга и сердца (рис. 5.8) [22, 23]. Поскольку в клетке есть ферменты, переводящие тестостерон в эстрадиол, неясно, происходит ли индукция благодаря тестостерону или эстрадиолу. Однако полученные результаты указывают путь управления уровнем специфических ферментов с помощью введения соответствующих количеств их индукторов.
Другим методом, который попытались применить для восстановления индукции ферментов у старых животных, был метод стимулирования регенерации ткани. Было показано, что активность митохондриальной малатдегидрогеназы понижается в печени молодых крыс после удаления надпочечника, но не меняется при этой же операции у старых животных [53]. Введение кортизона вызывает индукцию фермента у молодых крыс и не вызывает у старых. Однако если у старых крыс провести частичную гепатэктомию, дать печени возможность регенерировать в течение трех дней и затем ввести кортизон, то количество фермента увеличится. Таким образом, в отношении индукции митохондриальной малатдегидрогеназы регенерирующая печень старых крыс функционирует так же, как печень молодых. По-видимому, это нарушение индукции, связанное со старением, может быть ликвидировано. Наиболее значительным результатом этой работы является вывод о том, что молекулярные изменения, которые происходят в организме в процессе старения животного, могут быть обращены вспять. Однако клетки печени делятся на протяжении всей жизни, так как эти клетки являются премитотическими. Поэтому печень и способна к регенерации. Многие другие органы, такие, как мозг, сердце, скелетные мышцы, не могут регенерировать, так как их клетки теряют способность делиться на очень ранней стадии развития и становятся постмитотическими. Обратимы ли изменения, происходящие в старческом возрасте в этих органах?
В связи с упомянутыми исследованиями возникает еще один вопрос - обусловлено ли нарушение индукции малатдегидрогеназы в печени старых крыс уменьшением содержания специфических рецепторов кортизона или оно обусловлено репрессией гена малатдегидрогеназы? Поскольку активность малатдегидрогеназы в печени нормальных старых крыс высокая, появление репрессора в старческом возрасте, по-видимому, маловероятно. Чтобы ответить на поставленные вопросы, очевидно, важно измерить количество кортизон-специфического рецептора в печени старых крыс.
Описанные выше исследования показывают, что такие факторы, как гормоны и их рецепторы, важны для поддержания уровня и адаптивного ответа ферментов в различных возрастах. Изменения содержания этих факторов в старости могут вызывать изменения в транскрипции и трансляции и, таким образом, влиять на содержание ферментов. Это может в свою очередь приводить к функциональным сдвигам в органах и в организме в целом.
|
ПОИСК:
|
При использовании материалов проекта обязательна установка активной ссылки:
http://gelib.ru/ 'Геронтология и гериатрия'