Тепло и гены
Даже обычная температура тела животных является для существования их клеток отнюдь не безопасной. Про температуру около 37°С врачи говорят - она нормальная, биологи и физиологи - физиологическая. Но посмотрим, что происходит с молекулами при этой температуре.
О том, что некоторые ферменты при этой температуре спонтанно могут терять свою активность (инактивироваться), известно давно. Последствиям этой тепловой нестабильности белков серьезного значения не придавали, поскольку инактивированные молекулы белка в клетке могут быть заменены новыми. Но ведь генетическая информация таким образом не заменяется, хотя при определенных условиях имеющиеся искажения ее могут быть устранены.
В делящихся клетках она передается дочерним клеткам в результате процесса удвоения ДНК, ее редупликации. В неделящихся клетках, таких, как нейроны или мышечные клетки взрослых животных, редупликативного синтеза ДНК не происходит. Поэтому такие клетки должны функционировать в организме в течение всей его жизни с одним и тем же набором молекул ДНК. По крайней мере в течение первой ее половины должна сохраняться и генетическая информация женских половых клеток. Ведь гибель любой из этих клеток не может быть компенсирована делением оставшихся. Следовательно, в организме человека одна и та же ДНК находится во многих клетках при температуре 37° в течение многих десятков лет.
Теперь мы можем сформулировать вопрос, который, как это уже становится очевидным, является принципиальным для понимания молекулярно-генетических механизмов старения: с какой скоростью происходит тепловое повреждение (деградация) ДНК в клетке при 37°? Ответа на этот вопрос в первом издании этой книги мы дать не могли - к моменту ее написания еще не существовало достаточно точных методов определения скорости тепловой деградации ДНК при температурах, резко не отличающихся от физиологических.
В промежутке времени между двумя изданиями этой книги нам, а затем нескольким другим группам исследователей удалось разработать методики и с их помощью измерить скорость тепловой деградации ДНК в клетках человека и других млекопитающих при температурах, не намного превышающих 37°С.
Но сначала оценим приблизительно эту скорость из результатов опытов по тепловому повреждению (деструкции) ДНК в растворе. Установлено, что при такой деструкции "слабым местом" в ДНК является связь оснований с сахаром - так называемая гликозильная связь с ним пуринового основания. При нагревании растворов ДНК наблюдается прежде всего выщепление из ДНК пуриновых оснований - происходит процесс депуринизации ДНК. Это схематически показано на рис. 1.
После того как основание выщепилось из ДНК, связь фосфатной группы с сахаром (фосфодиэфирная связь) в участке депуринизации становится довольно неустойчивой, и она быстро подвергается разрушению вследствие присоединения молекулы воды (т. е. происходит гидролитический разрыв полинуклеотидной цепи).
Итак, в ДНК "спонтанно" протекают два основных повреждающих ее процесса - депуринизация и образование разрывов полинуклеотидной цепи. Скорости протекания обоих процессов зависят не только от температуры, но и от концентрации водородных ионов (рН) и ионов других солей (ионной силы раствора). Если хранить ДНК в водном растворе при значениях ионной силы и рН, близким к тем, при которых ДНК существует в клетке, то скорость депуринизации ДНК при "физиологической" температуре 37° будет столь мала, что ее трудно измерить с большой точностью современными методами количественного анализа пуриновых оснований. Однако можно еще определить скорость депуринизации ДНК и при других различных температурах и построить график зависимости константы скорости депуринизации от температуры. Этот график в так называемых Аррениусовых координатах имеет вид прямой (линия 1, рис. 5, где показана такая зависимость от температуры, линия 2 - скорости депуринизации лиофилизованной ДНК).
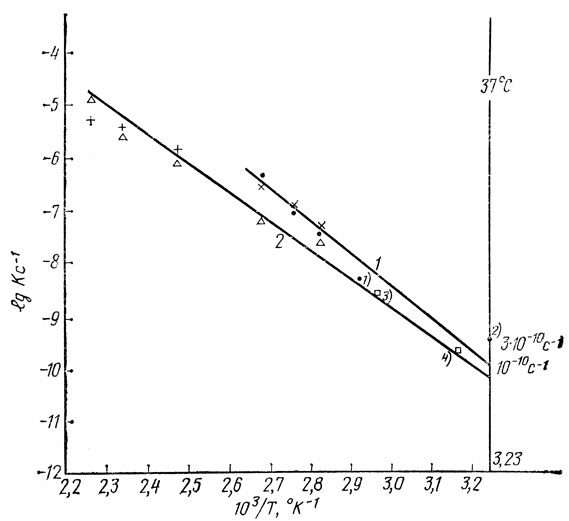
Рис. 5. Зависимость от температуры константы скорости депуринизации ДНК in vitro и образования суммы апуриновых участков и разрывов ДНК in vivo: I - выщепление аденина (·) и гуанина (x) из ДНК тимуса теленка в Na-цитратном буфере при значении ионной силы несколько большей физиологической и рН 6,8 и депуринизация ДНК бактерий (точка 1) или ДНК фага Т7 (точка 2) при значениях ионной силы и рН, близких к физиологическим условиям; точки 3 и 4 - образование тепловых повреждений ДНК in vivo соответственно, в клетках грызунов и в культивируемых фибробластах человека; II - выщепление аденина (Δ) или гуанина (+) из лиофильно высушенной ДНК тимуса теленка
Известные физико-химические свойства ДНК таковы, что кривую можно продолжить (экстраполировать) до значений абсциссы, соответствующей температуре 37°. Рассчитанная нами таким методом константа скорости депуринизации близка к значению 11-10 в секунду. Это означает, что из ДНК при физиологических условиях (т. е. при значениях температуры, рН и ионной силы, близких к таковым в клетках) каждую секунду в расчете на 1010 оснований должно теряться примерно одно основание.
Подробный теоретический анализ данных, касающихся организации ДНК в клетке и условий ее существования там, позволяет заключить, что выщепление пуриновых оснований из ДНК клетки происходит со скоростью, довольно близкой к той, которую мы рассчитали. Но даже если эта скорость в 10 раз меньше рассчитанной, заключение о вероятном значении в старении депуринизации ДНК при физиологической температуре остается тем же. Приходится только удивляться, каким образом клетки могут длительно существовать при температуре 37°.
Ответить на этот, казалось бы, весьма далекий от проблем "классической" геронтологии вопрос - значит, понять один из основных защитных молекулярных механизмов, выработанных, очевидно, уже на самых ранних этапах эволюции клеток и организмов для поддержания их жизнеспособности. А следовательно, найти подход для поиска факторов, увеличивающих устойчивость организма к старению.
Конечно, особенно интересны количественные данные теплового повреждения ДНК в клетке (in vivo). Определить скорость такого повреждения ДНК при 37°С до последнего времени было невозможно, так как при этой температуре наряду с возникновением тепловых повреждений ДНК протекает и обратный процесс - залечивания (репарации) этих повреждений (подробнее об этом процессе будет рассказано чуть позже).
Когда же попытались определить повреждения ДНК в клетках млекопитающих при температурах, превышающих 37°С, но меньше тех, при которых происходит разрушение клеток, никаких повреждений ДНК не зарегистрировали. Их обнаруживали лишь в случае прогревания клеток грызунов при 65°С. (Константа скорости образования повреждений ДНК при этой температуре была нами рассчитана, и ей соответствует точка 3 на рис. 5.) Таким образом, к началу 80-х годов среди биологов утвердилось мнение, что при температуре меньше 45°С тепловые повреждения ДНК в клетках млекопитающих если и индуцируется, то в количествах, которые невозможно зарегистрировать.
Однако с помощью усовершенствованного А. Н. Хохловым и мной седиментационного метода анализа ДНК в культивируемых фибробластах человека нам удалось измерить количество повреждений ДНК, индуцируемых в процессе прогревания этих клеток при 44°С. В эксперименте учитывались два существенных момента. Во-первых, в течение опыта клетки сохраняли жизнеспособность, т. е. можно было считать, что определяется процесс повреждения ДНК в живых клетках. И во-вторых, при температуре 44°С происходит существенное ингибирование процесса репарации ДНК, а это означает, что при такой температуре аккумулируются тепловые повреждения ДНК.
Рассчитанной на основании полученных нами данных константе скорости образования тепловых повреждений ДНК соответствует точка 4 на рис. 5.
Спустя несколько лет после опубликования этих данных ряд исследователей в США также зарегистрировали повреждения ДНК в клетках млекопитающих, прогретых при температурах меньше 45°С.
Несмотря на сложность анализа и в то же время его неполноту, можно с большой вероятностью утверждать: константа скорости теплового разрушения структуры ДНК лежит в пределах менее одного порядка, а именно в пределах 6·10-11 с-1 до 3·10-10 с-1.
Другие пути оценок скоростей спонтанного разрыва связей пуринов с сахарными остатками в ДНК, основанные на анализе механизмов разрывов этих связей, также приводят к значениям их констант при 37°С, лежащих в этом диапазоне. В том же диапазоне лежат и рассчитанные нами константы скоростей "спонтанного" разрыва полинуклеотидных цепей ДНК при физиологических условиях.
Но каков биологический смысл этих констант? Каким образом их знание приближает нас к пониманию механизмов старения? Без дополнительного количественного анализа ответить на эти вопросы невозможно.
Примерно каждую секунду в геноме каждой клетки человека спонтанно возникает одно повреждение ДНК. Чтобы "воспринять" приведенные константы, оценить значимость, казалось бы, ничтожно малых скоростей спонтанного возникновения в ДНК апуриновых участков и разрывов, рассчитаем число таких повреждений, возникающих в течение часа в геноме клетки человека и сравним эту величину с числом повреждений ДНК, индуцируемых в той же клетке ионизирующим излучением в дозе, вызывающей гибель значительной части облученных клеток. Такое сравнение ценно, и вот почему. Во-первых, основную "массу" повреждений ДНК, индуцируемых ионизирующим излучением, составляют также разрывы полинуклеотидных цепей или участки ДНК, из которых выщеплены пуриновые или пиримидиновые основания. Во-вторых, из многочисленных данных радиобиологии следует, что такого рода повреждения ДНК или их комбинации (в частности, образование из двух однонитевых разрывов, локализованных в комплементарных цепях вблизи друг друга, "полного" (двойного) разрыва двойной спирали ДНК) ответственны за биологические эффекты излучений. И в-третьих, содержание ДНК в нормальной клетке человека существенно не отличается от содержания в нормальных клетках других млекопитающих. Таким образом, наша количественная оценка скорости возникновения тепловых повреждений ДНК в клетках человека и ее значения будет справедлива и для клеток различных млекопитающих.
В геноме каждой клетки возникает одно повреждение ДНК в секунду. Согласно данным различных работ ДНК в каждой диплоидной клетке человека содержится около 7·109 пар оснований. Примерно столько же пар оснований содержится в ДНК соматической клетки других видов плацентарных млекопитающих. Таким образом, в расчете на ДНК диплоидной клетки человека скорость депуринизации можно принять соответствующей выщеплению в течение часа 2500 пуриновых оснований.
Поскольку процесс образования спонтанных однонитевых разрывов, как правило, лимитируется на этапе депуринизации ДНК, то, следовательно, скорость образования спонтанных однонитевых разрывов должна быть такого же порядка, что и скорость депуринизации ДНК. Подчеркнем однако, что сказанное верно лишь в отношении тепловых разрывов ДНК.
Но часть разрывов ДНК возникает в клетке вследствие реакции с ДНК эндогенных метаболитов радикалов ОН· и O 2, а также Н2О2, перекисей липидов ионов металлов с переменной валентностью. Кроме того, часть "спонтанных" разрывов ДНК обусловлена "ошибками" в работе ферментов (ДНКаз) и их "незапрограммированным" взаимодействием с ДНК. Таким образом, в зависимости от типа клеток и их физиологического состояния общая частота возникновения "спонтанных" разрывов в ДНК клеток человека, вероятно, в той или иной степени превышает значение 2·103 в час.
2, а также Н2О2, перекисей липидов ионов металлов с переменной валентностью. Кроме того, часть "спонтанных" разрывов ДНК обусловлена "ошибками" в работе ферментов (ДНКаз) и их "незапрограммированным" взаимодействием с ДНК. Таким образом, в зависимости от типа клеток и их физиологического состояния общая частота возникновения "спонтанных" разрывов в ДНК клеток человека, вероятно, в той или иной степени превышает значение 2·103 в час.
Количество повреждений ДНК такого типа, спонтанно возникающих в животных клетках в течение часа, примерно равно числу повреждений ДНК, индуцируемых в этих же клетках облучением их рентгеновским или γ-излучением в дозе 2,5-5 Гр (или 250-500 рад). Но при такой дозе облучения значительная часть клеток млекопитающих гибнет, а в организме этих животных обнаруживают изменения физиологических процессов.
Что касается скоростей возникновения других классов спонтанных повреждений ДНК in vivo, то определить их гораздо труднее. Межцепочечные сшивки в ДНК образуются в местах ее депуринизации, вероятно, с частотой в 10-2-10-3 от частоты депуринизации. Иными словами, в течение суток в геноме каждой диплоидной клетки человека образуется порядка 60-600 сшивок между обеими цепями ДНК. Скорость выщепления пиримидиновых оснований в расчете на геном диплоидной клетки человека, вероятно, примерно в 20 раз меньше скорости выщепления пуриновых оснований. Во всяком случае, соотношение скоростей выщепления пиримидиновых и пуриновых оснований из ДНК, находящейся в растворе, примерно таково. Следовательно, число пиримидиновых оснований, спонтанно выщепляемых из ДНК диплоидной клетки человека, может составлять около 3·103 за сутки. Это количество весьма существенное.
Сравнение на основании данных константы скорости спонтанного дезаминированного цитозина в растворе при рН 7 и при относительно высокой температуре с расчетным значением константы скорости депуринизации ДНК при той же температуре и том же рН показывает, что скорость спонтанного дезаминирования цитозина может быть даже больше, чем скорость депуринизации. Но последняя согласно приведенным выше расчетам соответствует выщеплению из ДНК в течение суток (в расчете на геном диплоидной клетки млекопитающих) порядка сотни тысяч аденина или гуанина. Следовательно, возможно, что скорость дезаминирования цитозина в ДНК составляет порядка 105 в течение суток в расчете на геном клетки млекопитающих. Если это так, рассматриваемое спонтанное изменение ДНК также нужно считать биологически очень существенным, учитывая резкое изменение матричных свойств цитозина после его дезаминирования.
В начале 80-х годов автор этой книги сформулировал предположение о том, что в клетке может протекать не программированный (спонтанный) процесс алкилирования ДНК в основном через реакцию ее с физиологическим донором метальных групп - S-аденозилметионином. Оценки показали, что в результате такой реакции только число метальных групп, спонтанно присоединенных к ДНК, составляет в расчете на геном клетки млекопитающих несколько сот в час.
В последние годы были проведены и экспериментальные исследования частоты непрограммированного метилирования ДНК, т. е. переноса на нее метальных групп с S-аденозилметионина. Из этих данных следует, что число метальных групп, спонтанно присоединяемых лишь по шестому атому кислорода гуанина в ДНК, составляет порядка 100 в час.
Теперь суммируем результаты проведенных оценок частоты возникновения различных спонтанных повреждений ДНК в клетках млекопитающих при физиологических условиях их существования. В течение часа выщепляется примерно 2500 пуриновых и 120 пиримидиновых оснований; индуцируется около 2000 однонитевых разрывов; дезаминируется значительное число цитозинов и метилируется, вероятно, не менее 100 гуанинов. Таким образом, общая скорость возникновения рассмотренных нами спонтанных повреждений ДНК составляет примерно 5·103 в час.
Это очень большое число повреждений ДНК, если принять во внимание биологическую роль ДНК. Ведь если в геноме клетки образуется за секунду по крайней мере одно повреждение ДНК, то за время жизни делящейся клетки (между двумя ее делениями в организме человека и многих других животных обычно проходит по меньшей мере 24 часа) в ней должно возникнуть 105 таких повреждений, а за время жизни нейронов в мозге человека (приблизительно 70 лет) - около 2,5·109 повреждений. Это соответствует выщеплению из генома большей части всех пуриновых оснований.
Частота повреждений ДНК, индуцируемых в каждой клетке организма млекопитающих за 1 час под влиянием фона ионизирующих излучений, составляет примерно 5·10-3. (Оценка сделана экстраполяцией зависимости от дозы числа индуцируемых повреждений ДНК различного типа к фоновым дозам.) Таким образом, скорость возникновения спонтанных повреждений ДНК примерно в 106 раз (а может быть, и в еще большее число раз - мы оценили "вклад" отнюдь не всех механизмов спонтанного повреждения ДНК) превышает скорость возникновения повреждений ДНК, индуцируемых фоновым излучением.
Это заключение может показаться парадоксальным. Ведь генетики, в том числе выдающиеся (например, академик Н. П. Дубинин), считают, что фон излучений вносит хотя и не очень большой, но все же измеримый вклад в частоту спонтанных мутаций у млекопитающих. Как же разрешить этот парадокс?
|
ПОИСК:
|
При использовании материалов проекта обязательна установка активной ссылки:
http://gelib.ru/ 'Геронтология и гериатрия'