Изменение функциональной деятельности нервной системы в процессе старения
Многочисленными исследованиями советских ученых установлено, что развитие человека - его рост, созревание и старение - происходит при непосредственном участии и контроле центральной нервной системы. Обнаружено, что у птиц и млекопитающих после удаления больших полушарий головного мозга наступают тяжелые нарушения сердечно-сосудистой системы, дыхания, желудочно-кишечного тракта, изменяется обмен веществ и энергии и т. д.
У децеребрированных животных наблюдается ослабление защитных свойств, появляется повышенная чувствительность к химическим и физическим воздействиям, к различным инфекционным заболеваниям, отмечаются трофические нарушения кожных покровов, изменения в различных органах и системах организма.
У молодых животных после удаления головного мозга появляются признаки преждевременного старения, которые часто приводят и к преждевременной смерти. М. К. Петрова (1944) показала, что не только удаление головного мозга, но и функциональное нарушение нормальной деятельности высших отделов нервной системы может привести к преждевременному старению и преждевременной смерти.
Нарушение динамики нервных процессов в высших отделах нервной системы отражается и на функциональной деятельности всего организма. Так, неврозы могут сопровождаться изменениями в деятельности различных систем - крови и кровообращения, пищеварения, обмена веществ, трофических функций, выделительных органов и т. д.
Некоторые исследователи полагают, что основной причиной старения и является изменение нервной системы, которое в дальнейшем ведет к нарушению функциональной деятельности организма в целом. Регулирующая роль центральной нервной системы в организме человека сохраняет свое ведущее значение во все возрастные периоды, а поэтому представляет большой интерес изучение происходящих изменений в самой центральной нервной системе в процессе старения.
Высшая нервная деятельность в процессе старения. Высшая нервная деятельность есть совокупность наиболее сложных форм работы коры больших полушарий и ближайших подкорковых образований, обеспечивающих приспособление организма животных и человека к окружающей среде. Вводя понятие "высшая нервная деятельность", И. П. Павлов оттенил ее в отличие от низшей нервной деятельности, которая регулирует и объединяет работу различных систем организма.
Основные реакции у животного и человека на все внешние и внутренние раздражения выражаются в форме рефлексов. Рефлекс - это сложная реакция организма, осуществляемая нервной системой в ответ на раздражение воспринимающих чувствительных окончаний. Рефлекторный акт - это ответная реакция организма на действие раздражителей. Такие явления, как кашель, сужение зрачков от яркого света, отдергивание конечностей при болевых раздражениях и т. п., являются врожденными, или так называемыми безусловными рефлексами.
Однако для существования высших животных одних врожденных рефлексов недостаточно. Для многообразных взаимоотношений организма с окружающей средой, для его быстрых и сложных реакций на внешние раздражители необходимо, чтобы некоторые раздражения действовали не только непосредственно, но и являлись сигналами на расстоянии.
Большая заслуга И. П. Павлова состоит в том, что он, изучая деятельность нервной системы, открыл качественно новую форму связи организма с окружающей средой - временные нервные связи, или так называемые условные рефлексы. Условные рефлексы образуются на основе безусловных рефлексов. Образование условного рефлекса происходит тогда, когда какой-либо безразличный для организма раздражитель, например свет или звонок, совпадает с действием безусловного раздражителя, например кормления.
Многократное сочетание звуковых сигналов (звонка) перед приемом пищи у животного ведет к образованию условного рефлекса, который проявится в том, что уже один звонок будет вызывать у животного обильное отделение слюны и соответствующую двигательную реакцию. Условные рефлексы вырабатываются, закрепляются и угасают в процессе жизни данного индивида. Они являются строго индивидуальными.
Если безусловные рефлексы вызываются ограниченным числом специфических раздражителей, условным раздражителем может являться всякое изменение в окружающей среде, воспринимаемое организмом. Условный рефлекс может образоваться и на прекращение воздействия какого-либо раздражителя, например на окончание звучания звонка. В данном случае возбудителем будет уже не появление, а, наоборот, исчезновение действующего фактора. В качестве условного раздражителя возможен и сильно действующий безусловный разрушительный раздражитель - прижигание кожи, ранение, электрический ток и т. д.
Условный рефлекс может быть выработан и на определенный отрезок времени. Так, если животному давать еду через строго определенные интервалы, то вскоре у него вырабатывается условный рефлекс и к моменту подачи пищи будет обильно выделяться слюна. Новый условный рефлекс можно выработать и на основе уже имеющегося сильного и прочного условного рефлекса. Тогда вновь вырабатываемый рефлекс будет называться условным рефлексом второго порядка.
Биологическое значение условных рефлексов заключается в том, что немногочисленные возбудители врожденных рефлексов при определенных условиях временно связываются с многочисленными и разнообразными явлениями окружающей среды. Благодаря временным нервным связям организм приобретает возможность быстро и точно реагировать на окружающую обстановку. При этом роль отдельных раздражителей может и изменяться. Они могут не только возбуждать, но и тормозить деятельность отдельных органов или систем, если это необходимо в интересах организма.
Следовательно, условные рефлексы могут быть положительными и отрицательными, тормозными. Положительные условные раздражители, вызывающие возбуждение, при определенных условиях могут превратиться в отрицательные и вызывать торможение. Внутреннее торможение развивается постепенно, иногда с большим трудом.
Образование и осуществление условных рефлексов свойственно высшим отделам головного мозга, коре больших полушарий. У животных условно-рефлекторная деятельность выполняется тем отделом центральной нервной системы, который является у них высшим. Так, у рыб и амфибий - промежуточным мозгом, у птиц - большим мозгом. Если у собаки удалить кору больших полушарий головного мозга, то основные реакции у нее сохранятся: животное будет избегать сильных раздражений, будет поднимать уши и голову при звуках и т. д. Но предоставленная сама себе собака сделается полным инвалидом, которую надо кормить, оберегать и т. п., иначе быстро погибнет, потому что в связи с операцией она потеряла то, что приобрела в процессе жизни - условные рефлексы. Выработать новые условные рефлексы, обучить чему-нибудь такое животное невозможно.
Изучение изменений функциональной деятельности высших отделов нервной системы при старении начато И. П. Павловым и его сотрудниками. Первые наблюдения, проведенные А. В. Тонких (1912) на собаках, показали, что ранее выработанные условные рефлексы у старых животных держались довольно прочно, новые же у них вырабатывались с большим трудом и характеризовались непрочностью и непостоянством. Автор объяснил это ослаблением процесса возбуждения в старческом возрасте.
В дальнейшем выяснилось, что расстройства тормозных процессов в старческом измененном мозгу имели еще более выраженный характер, чем нарушения процессов возбуждения. Выработать у старых животных дифференцировку было трудной задачей, несмотря на то, что сам дифференцировочный раздражитель затормаживал условный рефлекс почти до нуля.
Дифференцировка вырабатывается следующим образом: один раздражитель, например звонок, подкрепляется едой, а второй раздражитель, тоже звонок, но другой силы, не подкрепляется пищей. Вначале животное отвечает обильным выделением слюны и двигательной реакцией на оба раздражителя, но затем оно начинает различать эти звонки и отвечает только на тот, который подкрепляется едой. Таким образом, выясняется, что большие полушарии головного мозга различают, дифференцируют близкие раздражители.
У старых животных с большим трудом происходит и угашение выработанных условных рефлексов. Угашение, или угасательное торможение, происходит при повторении условных раздражителей без подкрепления.
Сопоставляя данные, полученные на различных животных, исследователи пришли к заключению, что функциональные старческие изменения коры головного мозга зависят также от типа нервной системы.
Как показали наблюдения, одни и те же раздражители у разных животных вызывают различную реакцию. У одних животных условный рефлекс вырабатывается быстро, у других - медленно. Одни животные легко переносят сильные внезапные раздражения, другие не выдерживают их. Анализируя эти особенности, И. П. Павлов пришел к выводу, что все разнообразие поведения человека и животных объясняется индивидуальной изменчивостью главных свойств нервной системы, их различными сочетаниями. Такими свойствами являются сила основных нервных процессов - раздражительного и тормозного, их равновесие и подвижность.
Исходя из степени преобладания возбудительного или тормозного процесса, из их взаимоотношений, установлено четыре типа нервной системы: 1) сильный, неуравновешенный с преобладанием возбуждения над торможением; 2) сильный, вполне уравновешенный, инертный, малоподвижный, медленный; 3) сильный, вполне уравновешенный, живой, подвижный тип; 4) слабый, со слабым возбуждением и торможением, легко тормозимый.
Наблюдая длительное время за собакой (10 лет), С. Н. Выржиковский (1935) обнаружил постепенное ослабление раздражительного и тормозного процессов, замедление условных рефлексов. Исследования показали, что подвижность нервных процессов у животных старческого возраста значительно снижена по сравнению с молодым.
На основании этих наблюдений ученые пришли к заключению, что функциональные свойства корковых клеток с возрастом изменяются. Понижается тонус коры больших полушарий. Это отражается как на возбудительном, так и на тормозном процессах.
М. К. Петрова на основании длительных экспериментальных исследований пришла к выводу, что в процессе старения организма ведущую роль играет центральная нервная система и главным образом кора головного мозга. Но нервная деятельность не есть изолированная деятельность мозговой коры, в тесной связи с нею функционируют вегетативная и эндокринная системы, а также мозжечок.
Характерной особенностью старых животных является то, что у них происходит замедление процессов восстановления. Если у молодых животных в интервале между раздражениями истраченный потенциал восстанавливается полностью, то у старых животных полного восстановления за это же время не происходит. А это значит, что новые раздражения падают на функционально нетрудоспособную клетку, что может привести к развитию хронического истощения и длительной функциональной неполноценности. Исходя из полученных данных, исследователи сделали выводы о том, что бороться со старостью надо не тогда, когда появились явные признаки ее, а гораздо раньше, чтобы не давать возможности им развиваться в молодом организме.
И. П. Павлов, подводя итоги наблюдений за старыми животными, пришел к заключению, что к старости системы условных рефлексов делаются неправильными, хаотическими. Животные к старости могут правильно работать только тогда, когда будет упрощена система рефлексов, уменьшено их количество. Он объяснял это явление снижением подвижности нервных процессов и их инертностью.
У человека условнорефлекторная деятельность при старческих изменениях нервной системы изучена еще не достаточно. Имеющаяся немногочисленная литература в основном посвящена изучению высшей нервной деятельности людей, страдающих тем или иным психическим заболеванием. Исследовательских работ, посвященных изучению организма здоровых стариков, совершенно мало.
Изучение условных рефлексов у людей в возрасте от 50 до 107 лет показало, что образование и упрочивание их в этом возрасте происходят медленно и с большими затруднениями. Обратило на себя внимание и непрочность рефлекса. Укрепившись к концу одного исследования, он ослабевал к началу следующего. С еще большим трудом происходит у них образование следового условного рефлекса и угашение. Если у молодых людей для угашения условного рефлекса требуется 8-10 применений неподкрепляемых условных раздражителей, то в старости для этих целей уже необходимо от 20 до 70 сочетаний.
Следовой условный рефлекс - это рефлекс, выработанный на след, т. е. на окончание раздражения. Если, например, рефлекс вырабатывается на звонок, то подкрепление дают не во время звонка, а тогда, когда прекратилось его звучание.
Дифференцировка у старых людей вырабатывается с большим трудом даже тогда, когда применяется большое количество дифференцировочного раздражителя.
Первые изменения высшей нервной деятельности у лиц старческого возраста наиболее четко проявляются при одновременном выполнении двух разных видов деятельности. Обнаружилось, что у молодых и среднего возраста людей ведущее значение имеет словесная система, в то время как при наступлении старости уже преобладает деятельность, основанная на безусловных рефлексах. У людей от 40 до 53 лет оборонительные условные рефлексы (мигательные) и двигательные (пищевые) осуществляются равномерно. У стариков же 67-84 лет, наоборот, наблюдалось или преобладание пищевых при умеренных возрастных изменениях высшей нервной деятельности, или оборонительных при глубоких старческих изменениях. При физиологической старости больше выражен пищевой рефлекс, в то время как при старческих патологических состояниях высшей нервной деятельности более выражены оборонительные реакции.
Обнаружено, что с возрастом одновременно с постепенным ослаблением основных нервных процессов усиливаются безусловные (пищевой и пассивно оборонительный) рефлексы, а ориентировочный рефлекс, т. е. рефлекс на обстановку, ослабевает. В более глубокой старости наблюдается быстрое истощение раздражительного процесса, который сменяется сонным торможением различной глубины. В результате ослабления регулирующей роли коры отмечаются изменения функциональной деятельности вегетативной нервной системы, что проявляется в нарушении правильности ритма дыхания и сосудистых реакций.
По данным некоторых исследователей, возбудительный и тормозной процессы, а также их подвижность усиливаются до 20 лет, затем они некоторое время стабильны, а с 30 лет снижаются. Начиная с 30 лет время двигательной реакции постепенно нарастает. Увеличивается и время скрытого периода реакции на положительный раздражитель вслед за применением тормозного раздражителя, что указывает на снижение подвижности нервных процессов. У старых людей в высших отделах нервной системы уже четко проявляется ослабление как возбудительного, так и тормозного процессов.
Исследования условно-рефлекторной деятельности практически здоровых людей (25 человек) в возрасте от 60 до 85 лет проводились и нами в секторе геронтологии АН БССР. Все испытуемые не имели каких-либо выраженных признаков заболеваний внутренних органов, чувствовали себя бодро, на состояние здоровья не жаловались. Не было у них и симптомов грубого поражения головного, спинного мозга и периферической нервной системы. Отсутствовали у них указания и на органические заболевания и повреждения нервной системы в прошлом. Все эти люди ведут активный образ жизни дома, многие из них выполняют общественную работу, участвуют в мероприятиях, проводимых горсоветом, следят за газетами, занимаются в кружках художественной самодеятельности (хоровом, драматическом).
Исследовали испытуемых всегда в утренние часы при хорошем их самочувствии. В случае малейших недомоганий или усталости опыты не проводились. На время исследования испытуемых помещали в затемненную камеру, относительно изолированную от шума. Опыты велись по речедвигательной методике А. Г. Иванова-Смоленского при одновременной записи биоэлектрической деятельности мозга. Сущность этой методики заключается в том, что при помощи словесного подкрепления у испытуемых вырабатывается условная связь между каким-либо индифферентным раздражителем (свет, звук) и ответной двигательной реакцией (испытуемый нажимал на электрокнопку). Время раздражения и ответная реакция испытуемого регистрировались на ленте электроэнцефалографа. Применялись ритмические световые и звуковые раздражители. Дифференцировкой служили той же силы свет и звук, только с другой частотой. Длительность раздражения 2 сек.
Исследование состояло из следующих этапов: образование положительной условной связи на световой раздражитель и дифференцировки к нему; образование положительной условной связи на звуковой раздражитель и дифференцировки к нему; переделка положительного условного рефлекса в отрицательный и отрицательного в положительный. Полученные результаты экспериментального исследования нейродинамики лиц пожилого возраста дают возможность сделать следующие выводы.
Образование условной связи на световые и звуковые раздражения у наших испытуемых не вызывало затруднения. Замыкание условной связи происходило на 2- 7-м сочетании. Связь эта относительно стойка и до применения дифференцировочного разражителя держалась прочно. Латентный (скрытый) период условного рефлекса у большинства испытуемых имел небольшие величины - 0,3-0,8 сек, только у двоих он составлял 1,2-1,5 сек. Однако величина латентного периода резко возрастала, как только начиналась выработка дифференцировки. При этом после применения дифференцировочного раздражителя с отрицательным подкреплением почти у всех испытуемых наблюдалось выпадение выработанного условного рефлекса.
Испытуемая Ч., 67 лет, в прошлом педагог (рис. 3). Условная связь на световой раздражитель с частотой 7 периодов в секунду у нее замкнулась уже на 2-м сочетании. Обращает внимание короткий латентный период 0,3 сек и его постоянство. Однако оно нарушалось, как только вводился дифференцировочный раздражитель (свет с частотой 12 периодов в секунду). После применения этого раздражителя с отрицательным подкреплением выпадал условный рефлекс. Следующее применение положительного раздражителя вызывало реакцию уже с большим скрытым периодом - 0,7 сек, применение дифференцировочного раздражителя во второй раз еще больше увеличило латентный период положительного условного рефлекса, который стал уже 1,2 сек, хотя дифференцировка еще не выработалась и испытуемая нажала кнопку отметчика так же, как и на положительный раздражитель. Увеличение латентного периода положительного условного рефлекса после дифференцировочного раздражителя есть результат взаимодействия со стороны второй сигнальной системы. После отрицательного подкрепления, хотя дифференцировка еще не выработалась, вызывалась уже тормозная реакция. Затем величина латентного периода на положительный условный раздражитель немного снизилась, однако была много выше, чем до применения дифференцировочного раздражителя. Дифференцировка выработалась на 4-м сочетании и уже была стойкой.
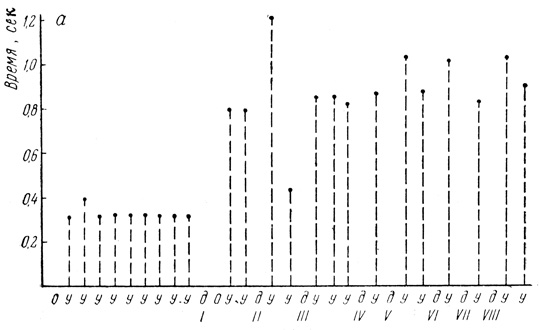
Рис. 3. Динамика величины латентного периода условного рефлекса на световой раздражитель (испытуемая Ч.) у - условный рефлекс; д - дифференцировочный раздражитель; о - основная реакция на приказ
Переделка положительного рефлекса в отрицательный, и наоборот никаких затруднений у большинства испытуемых не вызывала. У испытуемой Ч. отрицательный рефлекс стал положительным на 3-м сочетании, а положительный превратился в отрицательный на 2-м сочетании.
Испытуемая П., 68 лет, в прошлом бухгалтер (рис. 4). В качестве положительного условного раздражителя у нее был звук с частотой 6 в секунду, дифференцировкой служил той же силы звук с частотой 10 в секунду. Сила звука 70 дб. Звук передавался к креслу испытуемой через динамики. Условная связь на звуковой раздражитель замкнулась на 2-м сочетании. Латентный период был 0,4-0,5 сек. После применения первого дифференцировочного раздражителя два раза выпала условная связь. После вторичного его применения она выпадала трижды, несмотря на то что дифференцировка еще не была выработана. А последующая ответная реакция на положительный условный раздражитель была с очень большим латентным периодом (1,9-2 сек). В данном случае дифференцировка выработалась на 5-м сочетании. Здесь так же, как и у предыдущей испытуемой, было четко выражено последовательное торможение. Латентный период условной реакции после образования дифференцировки был в два с лишним раза больше по сравнению е исходным фоном. До начала выработки дифференцировки он равнялся 0,4-0,5 сек, а после его установления - 1,4-1,2 сек. До конца исследования латентный период оставался большим.
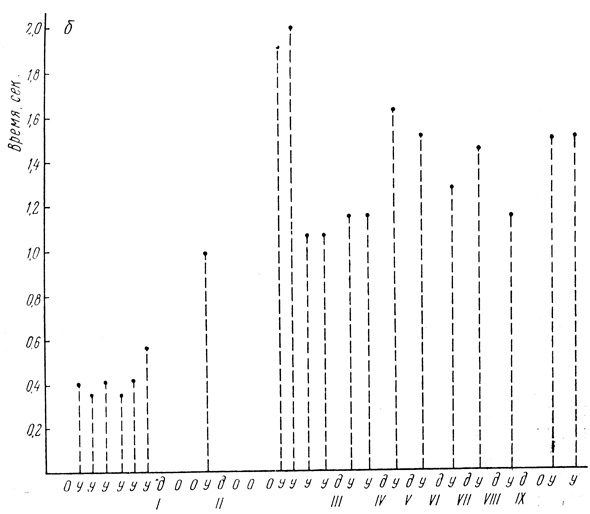
Рис. 4. Динамика величины латентного периода условного рефлекса на звуковой раздражитель (испытуемая П.). Обозначения те же, что и на рис. 3
Таким образом, условные рефлексы на ритмические световые и звуковые раздражения у наших испытуемых вырабатывались быстро, имели короткий латентный период и отличались относительной стойкостью. Образование дифференцировки к этим условным рефлексам также не представляло затруднения. Довольно легко произошла переделка условных раздражителей положительных в отрицательные и наоборот. Короче говоря, у испытуемых сила возбудительного процесса сохранена в достаточной степени. Тормозной процесс характеризовался некоторой инертностью, на что указывало четко выраженное последовательное торможение. После применения тормозного условного раздражителя с отрицательным подкреплением в большинстве случаев наблюдалось выпадение хорошо выработанного условного рефлекса и значительное увеличение латентного периода.
Изучение нейродинамических процессов в высших отделах нервной системы у человека нельзя считать полным без тщательного исследования функции второй сигнальной системы, т. е. нервных связей, свойственных только человеку. Для животных окружающий мир сигнализируется непосредственными раздражениями рецепторов и их следами в воспринимающих областях мозга. Эта первая сигнальная система. У человека, кроме этой сигнальной системы, имеется еще вторая сигнальная система. Она лежит в основе восприятия устной и письменной речи, речевой деятельности и словесного мышления. Рефлекторная, отражательная деятельность головного мозга человека является одновременно физиологической и психической. Физиологические явления далеко не однозначны по отношению к психическим, если рассматривать их в специфических для них свойствах и отношениях. Одно и то же явление может рассматриваться с различных аспектов. Одному и тому же физиологическому свойству могут соответствовать различные психические явления. Последние являются уже предметом изучения специальных наук - психологии и психиатрии, и в данной работе подробно касаться их мы не будем. Но поскольку психическая деятельность осуществляется мозгом, она подчиняется также закономерностям нейродинамики.
Слово является одним из наиболее эффективных условных раздражителей. Сила словесного воздействия как раздражителя объясняется его громадными ассоциативными связями с внешними и внутренними раздражениями благодаря предшествующей жизни взрослого человека. С появлением речевой сигнализации человек приобрел возможность анализировать и синтезировать бесчисленные сигналы нервной системы по новому принципу - принципу отвлечения и обобщения. Словесные раздражители дают возможность образовать многочисленные условно-безусловные и условно-условные временные связи. Речевая сигнализация развивается от более простых словесных сигналов до сложных отвлеченных представлений. Несмотря на то что первая и вторая сигнальные системы неразрывно связаны между собой, что их функции подчинены одним и тем же закономерностям, вторая сигнальная система имеет свои отличительные свойства. Качественное своеобразие деятельности второй сигнальной системы требует и дополнительных путей ее изучения.
Одним из приемов, получившим большое распространение для изучения сигнальных систем, является ассоциативный, или словесный, эксперимент. Суть его заключается в следующем: испытуемому предлагается набор слов в количестве 25. Слова обозначают конкретные предметы, окружающие данного человека в повседневной жизни, а также отвлеченные понятия, доступные пониманию независимо от уровня образования. Испытуемого инструктируют так, чтобы он на каждое слово-раздражитель давал ответы одним первым пришедшим в голову словом. Учитывались время ответной реакции, качество ответа, общая реакция испытуемого.
Этот метод позволяет изучить нейродинамические особенности преимущественно во второй сигнальной системе, уловить изменение корковой динамики от момента подачи словесного раздражителя до появления ответной словесной реакции. При этом не предполагается образование каких-то новых связей, а имеется в виду проследить динамику нервных процессов в системах, ранее образовавшихся и связанных с тем или иным предъявляемым словом.
Речевая сигнализация (письменная или устная) также проходит через онтогенетические ступени развития от более простых словесных сигналов (название предметов, действий) до сложных отвлеченных представлений. Все корково-подкорковые нейродинамические связи находятся в строгой субординации и взаимозависимости.
Выяснилось, что возрастные изменения в высших отделах нервной системы касаются и речевой деятельности. У испытуемых, особенно более позднего возраста, обнаружилось затруднение давать ответную реакцию одним словом согласно инструкции. В ряде случаев на одно слово-раздражитель они давали ответы целыми фразами. Это объясняется ослаблением внутреннего торможения.
В своей основе ответная реакция была правильной. У некоторых испытуемых к концу обследования отмечалось много повторных ответов. В отдельных случаях наблюдалась эхолалическая реакция - прежде чем дать ответ, испытуемый повторял слово-раздражитель по нескольку раз. Латентный период ответной реакции у стариков характеризовался непостоянством и большой его величиной по сравнению с молодыми людьми.
Исследование условно-речевых связей на одни и те же слова с различными интервалами показало, что, несмотря на большие промежутки времени между опытами, испытуемые на слова-раздражители дают одинаковые ответы. Это указывает на некоторую заторможенность второй сигнальной системы, на ограничение запаса слов. Наряду с этим нами были исследованы условно-речевые реакции на избранные раздражители. Этот эксперимент мы проводили в двух вариантах: в одних случаях испытуемым предлагали не отвечать на слова, обозначающие одушевленные или неодушевленные предметы; в других - отвечать только словами, противоположными по смыслу (большой - маленький, старый - молодой). Этот эксперимент требовал дифференциации раздражителей, потому что наряду с положительными реакциями включались и тормозные, когда испытуемые должны были задерживать ответ. Предлагая им давать ответ противоположного смысла, мы усложняли их задание по сравнению с предыдущим. Здесь уже была необходима достаточная сила активного, дифференцировочного торможения во второй сигнальной системе, достаточная концентрация возбудительного и тормозного процесса. Цель этих исследований - выявить взаимодействие тормозного и возбудительного процессов главным образом во второй сигнальной системе, их подвижность, силу внутреннего торможения.
В большинстве опытов испытуемые свои задания выполняли правильно. В некоторых исследованиях, несмотря на тщательную инструкцию с примерами, смысловое ограничение не принималось во внимание. Наконец, в ряде случаев задание выполнялось частично: после нескольких правильных реакций испытуемый давал неправильные ответы (происходило растормаживание), т. е. отвечал на те слова, на которые не должен был давать ответы.
На основании исследований функциональной деятельности сигнальных систем в старческом и молодом возрасте можно сказать, что в ходе старения в ряде случаев отмечается снижение силы активного торможения, что особенно четко проявляется в эксперименте со словесным ограничением. Наблюдается также инертность нервных процессов, показателем которой является большой латентный период ответной реакции. Заторможенностью второй сигнальной системы объясняется и бедность ассоциаций наших испытуемых.
Можно предположить, что ослабление силы и подвижности нервных процессов лежит в основе некоторых особенностей старческой психики. К ним прежде всего нужно отнести некоторое снижение психической активности, снижение способности усвоения нового материала, ослабление активной памяти и т. д. Снижение памяти у стариков, по мнению исследователей, следует отнести за счет некоторого общего снижения функциональной деятельности головного мозга, ослабления возбудимости нервных клеток и как следствие этого - ослабление раздражительного и тормозного процессов, их инертность. Однако нарушение памяти хотя и есть частое явление, но оно не обязательный компонент старческого возраста, ибо многие старики обладают относительно хорошей памятью. Память снижена обычно на текущие события, имена, даты. События далекого прошлого, как правило, в памяти сохраняются в достаточной степени даже у стариков свыше 90 лет. Рассеянность, свойственная старческому возрасту, некоторыми исследователями объясняется развитием в коре головного мозга так называемых отрицательных индукций. Пожилой человек, будучи сосредоточен на чем-либо или занят каким-либо делом, полностью отдается ему, и все остальное выпадает из его внимания. Один очаг возбуждения в коре мозга в силу закона индукционных взаимоотношений тормозит все другие пункты. Излишняя разговорчивость, которая отмечена у ряда стариков, может быть объяснена ослаблением торможения и низкой возбудимостью.
Как показали наблюдения, изменения высшей нервной деятельности происходят не параллельно возрасту. Следовательно, изменения функциональной деятельности высших отделов нервной системы, которые наблюдаются в старческом возрасте, нельзя объяснить только старостью. Необходимо учитывать и другие факторы, а именно: общее состояние организма в момент исследования, перенесенные заболевания, особенно заболевания центральной нервной системы, предшествующую трудовую деятельность, заинтересованность, понимание важности проводимых исследований испытуемым и т. д. Большую роль играет и тип нервной системы.
Многочисленные наблюдения показывают, что у людей, живущих интересами коллектива, занимающихся активно умственным и физическим трудом, функциональная деятельность высших отделов нервной системы продолжает оставаться на высоком уровне до глубокой старости. Тонус коры больших полушарий головного мозга сохраняется. Предполагают, что из всех систем целостного организма высших животных и человека наиболее устойчивой, пластичной, активно функционирующей и долгоживущей является система больших полушарий мозга.
В литературе есть немало примеров, которые говорят о том, что у многих людей только во вторую половину жизни полностью проявляются творческие способности, но для этого очень важно, чтобы человек до конца своей жизни не прекращал как умственной, так и физической работы. А по мнению известного румынского ученого К. Пархона, деятельность коры головного мозга оказывает тормозящее влияние и на процесс старения.
Биоэлектрическая деятельность мозга в процессе старения. Установлено, что почти все явления жизнедеятельности организма: движение мускулатуры, работа внутренних органов - сердца, желудка и др., а также активность высших отделов нервной системы - сопровождаются электрическими процессами. В течение всей жизни человека в высших отделах нервной системы протекают электрические процессы, которые являются показателями функциональной деятельности мозга. Они могут быть зарегистрированы специальными приборами - электроэнцефалографами. Обмен веществ мозга может ускоряться или замедляться во времени, увеличиваться или уменьшаться по своей интенсивности. В электроэнцефалограмме в какой-то степени и будет отражаться метаболизм нервных клеток.
Однако электроэнцефалограмма выражает только электрохимическую сторону обмена, в то время как тепловая, химическая, механическая и другие, составляющие обмен, не обнаруживаются методом электроэнцефалографии. Первые записи биоэлектрической деятельности мозга были сделаны Бергером в 1924 г. Он установил, что электрические колебания возникают в нейронной ткани. Они изменяются с возрастом, при чувствительных раздражениях, а также при физико-химических изменениях в организме. Применяемые обычно способы отведения биотоков мозга с поверхности черепа позволяют зарегистрировать суммарные эффекты биоэлектрических процессов больших групп нервных клеток в зоне отведения. Основной величиной, в наибольшей степени отражающей функциональное состояние коры мозга, является частота ритмов электроэнцефалограммы.
К сожалению, в электрофизиологии до настоящего времени нет общепризнанной теории происхождения ритмов электрических колебаний мозга. Однако установлено, что ритмические биоэлектрические потенциалы коры находятся в зависимости от уровня обмена веществ микроструктуры элементов коры головного мозга, а также от морфофизиологических связей коры с другими отделами головного мозга. С разных областей мозга регистрируются различные типы ритмов. Электроэнцефалограмма здорового человека, находящегося в спокойном состоянии, характеризуется относительно устойчивым ритмом.
Биоэлектрическая деятельность мозга здорового человека представлена в основном волнами двух родов: альфа-волнами и бета-волнами. Альфа-волны имеют частоту колебаний от 7,5 до 13 в секунду, амплитуду от 10 до 100 мкв. Они наиболее выражены в затылочной области. Бета-волны имеют в 2-4 раза меньшую амплитуду по сравнению с альфа-волнами. Частота их варьирует от 14 до 35 в секунду. Они локализируются преимущественно в прицентральных областях коры. При подавлении альфа-активности амплитуда бета-волн несколько возрастает. Значительно реже встречаются и другие виды волн. Быстрые колебания с частотой от 35 до 55 в секунду очень маленькой амплитуды - гамма-волны. Чаще всего они наблюдаются в височных областях. "Пики", или "иглоподобные" волны, встречаются при патологических состояниях. Амплитуда их колебаний может доходить до 150-200 мкв. Они могут быть одиночными, а также возникать группами. Встречаются и медленные волны с частотой ниже 7,5 гц в секунду (дельта-ритм с частотой от 0,5-3 и тэта-ритм от 4-7 гц). Они наблюдаются чаще во время сна и распространены в детском возрасте.
Однако при оценке функционального состояния коры головного мозга только на основании статической регистрации фоновых колебаний возникают большие затруднения. Известно, что в ряде случаев электроэнцефалограммы, записанные при физическом и умственном покое, у лиц с четко выраженными клиническими проявлениями заболевания мозга не обнаруживали нарушения биоэлектрической деятельности. Отсюда следует, что для того, чтобы выявить то или иное функциональное нарушение мозга, необходимо наблюдать его в деятельном состоянии. За последнее время широкое применение получило исследование биоэлектрической деятельности мозга при ритмических световых раздражениях.
Афферентные раздражения вызывают изменения электрической активности во всех областях коры, но они различны по своему характеру. Эти различия объясняются особенностями архитектонических полей отдельных областей коры мозга, связями исследуемой области с другими областями коры и подкорковыми образованиями, а также функциональным состоянием в момент исследования.
Установлено, что лучше всего альфа-ритм выражен в состоянии покоя и в состояниях, переходных от бодрствования ко сну (рис. 5). Все раздражающие воздействия (свет, звук, температурные раздражители и т. д., а также умственное напряжение) ведут к торможению, исчезновению альфа-ритма и к повышению амплитуды и частот бета-ритма.
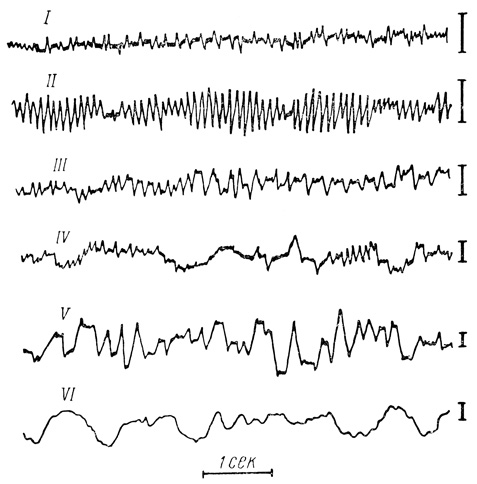
Рис. 5. Типичные электроэнцефалограммы, записанные при различных уровнях сознания: I - возбуждение; II - покой; III - дремотное состояние; IV - засыпание; V - глубокий сон; VI - кома. Калибровка 50 мкв (по У. Пенфилду, Г. Джасперу)
Одни авторы рассматривают угнетение альфа-ритма как проявление процесса торможения, другие, наоборот, связывают это явление с возбуждением в коре и подкорковых образованиях. В настоящее время бесспорным, очевидно, можно признать только то, что альфа-ритм является в какой-то мере показателем взаимоотношений процессов возбуждения и торможения, подвижности нервных процессов в высших отделах нервной системы. Следовательно, по их появлению и исчезновению косвенно можно судить о подвижности нервных процессов в высших отделах нервной системы. Длительных исследований биоэлектрической деятельности мозга стариков, как показывают литературные данные, до сих пор никем не проводилось. Обследуя биоэлектрическую деятельность мозга стариков в возрасте от 60 до 90 и больше лет, исследователи пришли к заключению, что установить четкой закономерности между изменениями электрической активности головного мозга и степенью одряхления организма не представляется возможным. Большинство исследователей считают, что после 60-летнего возраста наблюдается тенденция мозговых ритмов к замедлению. Может иметь место и смешанный тип электроэнцефалограммы с наличием медленных и частых ритмов. В ряде случаев наблюдается дезорганизация основного ритма, снижение корковой активности, иногда до полной сглаженности электроэнцефалографической кривой. Снижение корковой активности может также проявляться в уменьшении выраженности альфа-ритма, в увеличении латентного времени "угнетения" его на действие раздражителей. У большинства пожилых лиц обнаружено снижение частоты и выраженности альфа-ритма и увеличение медленных волн в электроэнцефалограммах. Уменьшение амплитуды с возрастом необязательно. Наоборот, в ряде случаев отмечается даже увеличение ее.
Обращает внимание широкий диапазон индивидуальных различий. Некоторые исследователи полагают, что чем старше возраст испытуемых, тем чаще встречается медленный доминирующий ритм, тем чаще бывают отклонения. Но человек преклонного возраста может и не иметь никаких отклонений. Поэтому другие авторы считают, что электроэнцефалограмма стариков лишена каких-либо особенностей, за исключением некоторого замедления волн альфа-ритма. Выраженность изменений электрической активности головного мозга определяется прежде всего наличием патологического, атеросклеротического процесса, а не возрастом. При отсутствий сосудистых нарушений и сохранности работоспособности альфа-ритм хорошо выражен, нарушение сердечной деятельности может вызвать патологию и при отсутствии расстройств мозгового кровообращения.
Изменения электроэнцефалограммы стариков чаще диффузны, двусторонни, но могут быть и локализованными, преимущественно в лобных или височных областях. Отмечено снижение реакции перестройки ритмов мозга на ритмические раздражители (световые, звуковые).
В исследованиях, проводимых нами, биоэлектрическая деятельность мозга регистрировалась многократно электроэнцефалографом. Запись биоэлектрической деятельности велась при моно- и биполярном отведениях с различных участков полушария в затемненной, относительно звукоизолированной комнате, в покое, при закрытых глазах, а затем при ритмических световых и звуковых раздражениях. Последние производились фонофотостимулятором.
Ритмические световые раздражения давались с частотой от 1 до 50 мельканий в секунду. Сила света была постоянной. Расходуемая при каждой вспышке энергия 0,2 Дж. Частота и длительность раздражения регистрировались на ленте электроэнцефалографа. Раздражения в каждом опыте давались по-разному - с возрастающей частотой от 1 до 50 мельканий в секунду и, наоборот, с постепенным снижением от высоких частот к низким. В некоторых опытах различная частота была с большими интервалами, отдыхом. Длительность раздражения также варьировала от 1 сек до 1 мин и более. В тех случаях, когда испытуемые не выдерживали яркости света и часто мигали, раздражения давались при закрытых глазах или через фильтры различных цветов.
Анализируя полученные данные по исследованию биоэлектрической деятельности мозга пожилых людей в покое и сравнивая их с ранее проведенными нами наблюдениями на группе здоровых молодых людей в возрасте от 18 до 35 лет (1961), мы обнаружили довольно четкие различия между ними. Прежде всего у пожилых людей снижается частота альфа-ритма. Если у молодых людей она может достигать 12, а иногда и 13 Гц в секунду, то у наших испытуемых она равнялась 8-9 в секунду. Только у двоих частота альфа-ритма была 10-11 в секунду.
У некоторых испытуемых наблюдалось уменьшение и времени альфа-ритма, доходившее до 12-30%. В ряде случаев удалось обнаружить и другие виды волн, как-то: тета-, дельта-волны, сдвоенные альфа-волны, веретенообразные. Но эти волны наблюдались только изредка, в отдельных областях, чаще в теменных и височных и не у всех испытуемых.
Еще в 1934 г. Эдриан и Меттюз обнаружили, что при раздражении сетчатки глаза ритмическими световыми раздражениями происходит перестройка корковой активности затылочной области в соответствии с частотой подаваемых раздражений. В дальнейшем эти данные были подтверждены многочисленными исследователями. Выяснилось, что у большинства здоровых людей биоэлектрическая деятельность мозга изменяется в ритме раздражений в пределах от 1 до 30 миганий в секунду. Верхний предел усвоения ритма 12-30 миганий в секунду, нижний - в области 2-8 в секунду. Согласно данным некоторых авторов, строгое согласование местных корковых ответов на ритмичное световое раздражение наблюдается иногда и при частоте 60 миганий в секунду.
Исследование биоэлектрической деятельности мозга при ритмических световых раздражениях у наших испытуемых также показало большую разницу по сравнению с молодыми людьми. Выяснилось, что диапазон усвоения ритма у пожилых людей значительно сужен и сдвигается больше в сторону редких частот. В большинстве случаев у испытуемых диапазон усвоения находился в пределах 2-15 колебаний в секунду. Иногда усвоение было еще более ограниченным от 5 до 10, от 4 до 8 колебаний в секунду и т. д. У некоторых испытуемых оно было нечетко выражено или совсем отсутствовало. Так, у испытуемой П., 68 лет (рис. 6), запись биоэлектрической деятельности мозга до раздражения показала преобладание альфа-ритма с частотой 9-10 в секунду, амплитуда 30-60 мкв. Альфа-ритм лучше всего выражен в затылочных областях. Отмечается симметричность межполушарных взаимоотношений.
При нанесении ритмических световых раздражений возрастающей частоты на сетчатку глаз обнаруживается исчезновение альфа-ритма и перестройка биоэлектрической деятельности в ритме наносимых раздражений. Как видно из рисунка, диапазон усвоения у данной испытуемой от 3 до 12 мельканий в секунду. Раздражения, наносимые с большей частотой (15 мельканий и выше), мозгом данной испытуемой не усваиваются.
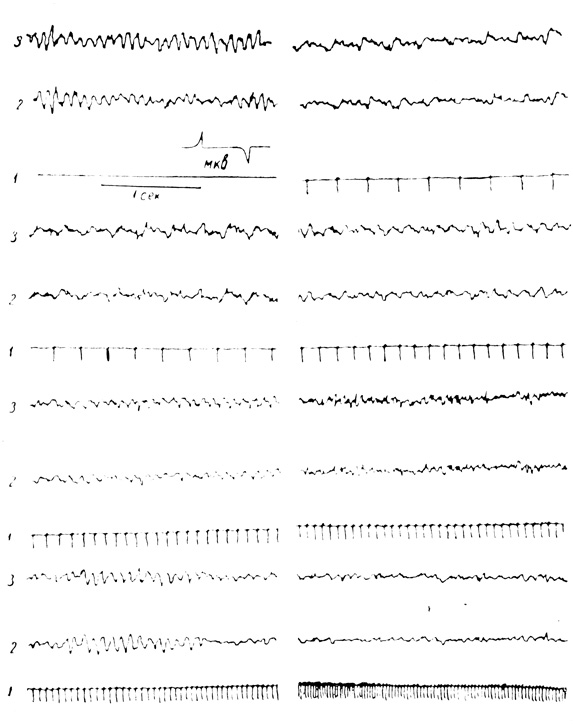
Рис. 6. Электроцефалографическая кривая усвоения ритма световых раздражителей (испытуемая П.): 1 - отметка световых раздражений; 2 - затылочная область слева; 3 - затылочная область справа
Учитывая, что критерием функциональной подвижности ткани, согласно учению Н.Е. Введенского (1952), является то наибольшее количество электрических колебаний, которое физиологический субстрат может воспроизвести в единицу времени, оставаясь в точном соответствии с ритмом наносимых раздражений, можно считать, что показание усвоения ритма световых раздражений и является критерием функциональной подвижности мозга (в данном случае коркового отдела зрительного анализатора). Поэтому усвоение ритма световых раздражений высокой частоты можно считать показателем высокой подвижности (лабильности) мозга, в то время как отсутствие усвоения или усвоение только небольшого диапазона частот является показателем пониженной лабильности. По сравнению с молодыми людьми у наших испытуемых наблюдается четко выраженное снижение функциональной подвижности мозга.
Амплитуда электрических колебаний при световых раздражениях заметно снижалась по сравнению с исходным уровнем. Следовательно, реакции коры на световые раздражения относятся к тормозному типу. Что касается звуковых раздражителей, то они подавались также с частотой от 1 до 50 в секунду, силой 70-75 Дб. Четкого усвоения ритма звуковых раздражений нам отметить не удалось. Имеются отдельные участки мозга, где как бы намечается перестройка, но она никогда не была точной в ритме подаваемых раздражений. Чаще всего можно было наблюдать только изменение амплитуды в сторону увеличения или уменьшения и некоторую перестройку ритма, которая у каждого испытуемого была выражена по-разному. Определенных закономерностей в динамике биоэлектрической деятельности на звуковые раздражения выявить не удалось.
По сравнению с молодыми людьми у наших испытуемых наблюдается некоторая инертность нервных процессов в высших отделах нервной системы, на что указывает более ограниченный диапазон усвоения ритма световых раздражений, сдвиг его к редким частотам. Функциональные изменения высшей нервной деятельности являются следствием морфологических и биохимических изменений в ней.
Изучение морфологических изменений в центральной нервной системе показывает противоречивые результаты. Одни исследователи находят ряд изменений мозга, которые склонны связывать со старческим возрастом. Другие отрицают существенные изменения мозга в связи с возрастом, полагая, что нет основания считать, что неповрежденная и сохранившая нормальное питание нервная система будет функционировать иначе, чем в молодости, только потому, что прожито много лет. Тем не менее некоторые изменения функциональной деятельности нервной системы все же имеются. С возрастом развиваются процессы старения и в самой нервной системе.
За старческими изменениями в центральной нервной системе велись наблюдения еще в глубокой древности. На изменение объема мозга в процессе старения указывали еще Аристотель, Гиппократ, Гален и др. Давно отметили, что извилины мозга стариков сглажены, вещество мозга мягче по консистенции, мозговые оболочки утолщены, в желудочках обнаружено скопление жидкости и т. д.
Вес мозга в течение жизни изменяется. У мужчин рост мозга продолжается до 25 лет, у женщин достигает своей максимальной величины к 20 годам. Затем некоторое время вес мозга держится на одном уровне, а примерно с 50 лет начинает заметно уменьшаться. Согласно данным исследователей, у мужчин вес мозга меньше поддается процессам уменьшения по сравнению с женщинами. Морфологически процесс старческого изменения мозга выражается в дегенеративных и атрофических явлениях. В процессе старения постепенно происходит уменьшение количества нервной ткани мозга и замена ее грубыми коллагеновыми волокнами, волокнистой глией, фиброзными астроцитами.
Изменения нервных элементов мозга к глубокой старости могут выражаться в форме нарушения ядерно-плазматических отношений, уменьшения, сморщивания ганглиозных нервных клеток и вакуолизации их, гиалиновой дегенерации, гидропическом расширении клетки, сморщивании и частичном растворении хромидальных веществ и т. д. В ряде случаев наблюдается и уменьшение ядрышка. Может иметь место гипертрофия и отпочкование дендритов. У старых животных отмечено и разрыхление внутриклеточной фибриллярной сети.
Типичным для позднего возраста является отложение так называемой старческой пигментации, накопление желто-коричневых липоидных и черных меланоидных пигментов. Есть предположение, что образование пигмента связано с нарушением обмена в клетке. Как показали дальнейшие исследования, пигментирование начинается уже в раннем возрасте и к старческому периоду достигает выраженной степени. Однако в молодом возрасте пигментные зерна окрашиваются в бледно-желтую окраску, а в более позднем они приобретают темно-коричневую окраску. Наиболее богаты "старческим пигментом", или липофусцином, клетки симпатических узлов. В ряде случаев пигмент покрывает все структурные образования клетки. Но не все клетки мозга и нервных ганглиев стариков насыщены пигментами. Есть клетки совершенно свободные от старческой пигментации.
Процесс старения отражается и на периферических нервах. Обнаружено, что с возрастом постепенно понижается скорость проведения возбуждения по нервам. Так, нашли, что скорость проведения импульса по локтевому нерву в возрасте от 20 до 29 лет равняется 57,9 м/сек; от 30 до 39 лет - 57,7; от 40 до 49 - 56,6; от 50 до 59 лет она снижается до 53,2 м/сек. Морфологические изменения в нервах наиболее часто выражаются в жировой инфильтрации их, уменьшении плотности нервных волокон в его проводящей системе.
Изменения в различных отделах нервной системы выражены не в одинаковой степени. В одних отделах процесс выражен четко, в то время как в других он или совсем не обнаруживается, или имеется в слабой степени. Некоторые связывают это с кровоснабжением, полагая, что больше страдают участки с переродившимися кровеносными сосудами. И действительно, значительная часть изменений мозга может быть отнесена за счет сосудистых нарушений.
Сосудистые поражения головного мозга стариков касаются как крупных, так и мелких сосудов, как внутренней, так и средней и наружной их оболочек.
Однако момент появления сосудистых нарушений определить трудно, так как они возникают постепенно, в различные периоды жизни, часто уже тогда, когда еще нет никаких признаков старости. Поэтому до сих пор не установлены характерные для старческого возраста сосудистые изменения головного мозга. Ранее существовавшее мнение об артериосклеротических изменениях как типичных для старости в настоящее время отвергнуто, так как этот процесс может быть четко выявлен и у молодых и не быть в глубокой старости.
Изменения, наблюдающиеся в сосудах мозга в процессе старения, сводятся к атрофии артериальной стенки, уменьшению мышечного слоя и эластической ткани. Сосуды становятся гипотоничными, теряют способность сокращаться и частично или полностью утрачивать свою сосудодвигательную функцию. Отмечается и атрофия капилляров с уменьшением сосудистого калибра до 1/4 первоначального размера.
В ряде случаев кровеносные сосуды головного мозга у стариков находятся в виде регидных трубок. Изменения сосудов мозга ведут к нарушению кровообращения, а следовательно, и питания мозга. Процесс старения кровеносных сосудов мозга находится в зависимости от многих факторов внешней и внутренней среды. Развитие процессов старения в самой нервной системе неизбежно накладывает отпечаток на жизнедеятельность всего организма.
Повышенная или пониженная возбудимость, которая развивается под влиянием тех или иных воздействий у молодых, быстро уступает место полному восстановлению прежнего уровня. В пожилом возрасте это восстановление происходит более замедленными темпами.
Снижение силы и подвижности нервных процессов в высших отделах нервной системы отражается и на функциональной деятельности организма. Высшие отделы нервной системы не смогут, как в молодости, быстро реагировать на информирующие сигналы с различных областей организма и не в состоянии быстро посылать стимулирующие и задерживающие деятельность импульсы. Снижается лабильность высших отделов нервной системы в отношении управления процессами организма. В связи с этим могут возникать различные дисгармонии в деятельности отдельных органов и систем организма, чувствительность которых к нервным импульсам в процессе старения снижается. Возникает качественно новый уровень деятельности организма, новый приспособительный механизм, который проявляется в повышении чувствительности к гуморальным факторам. Следовательно, изменяется характер влияния высших, регулирующих деятельность организма отделов к объектам регуляции. Однако, как и прежде, высшие отделы нервной системы остаются ведущими и регулируют все явления, происходящие в организме.
|
ПОИСК:
|
При использовании материалов проекта обязательна установка активной ссылки:
http://gelib.ru/ 'Геронтология и гериатрия'