5.1. Программа самоликвидации или износ?
При обсуждении гипотезы запрограммированности продолжительности жизни ее сторонники в качестве основного аргумента приводят обычно факт больших межвидовых различий по срокам жизни [Комфорт, 1967; Comfort, 1979]. Этот на первый взгляд убедительный аргумент на самом деле ничего не доказывает и свидетельствует лишь о необходимости уяснения смысла понятия "программа". Действительно, разные технические устройства также могут иметь совершенно разные характерные "сроки жизни" (сравните, например, сроки службы электрической батарейки и фотоэлемента). Огромные различия наблюдаются также во времени жизни атомов разных химических элементов и их изотопов. Было бы, однако, опрометчиво на основании только этих фактов делать вывод о существовании внутри каждого атома специальной программы, ограничивающей срок его жизни. Представляется более разумным, что время жизни систем (включая биологические) определяется просто особенностями их устройства и функционирования. Таким образом, факт больших межвидовых различий по продолжительности жизни свидетельствует лишь о том, что ее величина (так же как и величина всех остальных признаков) определяется материальной природой изучаемых объектов. Другой аргумент, который обычно приводят сторонники гипотезы запрограммированности продолжительности жизни, связан с утверждением о высокой наследуемости сроков жизни. Этот аргумент содержит сразу две ошибки. Во-первых, как мы убедились на основании приведенных ранее данных (см. раздел 2.3), наследуемость сроков жизни (в смысле сходства между кровными родственниками) необычайно низка. Представление о высокой наследуемости продолжительности жизни отчасти связано с тем, что иногда не различают понятия достоверности влияния и силы этого влияния. В результате, установив безусловную справедливость факта влияния длительности жизни родителей на продолжительность жизни потомства, делают ошибочный вывод о безусловной генетической детерминированности этого признака. Во-вторых, высокая наследуемость продолжительности жизни является необходимым, но не достаточным условием для того, чтобы считать этот признак генетически запрограммированным. Так, например, высокие гастрономические качества осетровых рыб. несомненно, генетически детерминированы, однако из этого вовсе не следует, что в их геноме существует специальная программа, рассчитанная на удовлетворение вкусов человека. Таким образом, мы приходим к необходимости уточнения смысла понятия "программа", несмотря на его кажущуюся простоту.
Размышления о смысле понятия "программа" приводят нас к выводу о том. что это понятие предполагает существование определенной цели, достижению которой она служит. В этом смысле можно говорить, что многие полезные признаки сельскохозяйственных животных и растений генетически запрограммированы, поскольку в отношении этих признаков долгое время велся целенаправленный отбор. В случае же естественных условий жизни выбор цели определяется борьбой за существование и отбором по тем признакам и в том конкретном направлении, которые способствуют выживанию в данных конкретных условиях. Таким образом, о запрограммированности признака можно говорить только тогда, когда существует понятие цели и можно ответить, кем (человек) или чем (эволюция) такая программа была сформирована. Следовательно, вопрос о запрограммированности продолжительности жизни сводится к выяснению ее значения для биологической эволюции.
Если с эволюционных позиций подойти к проблеме запрограммированности продолжительности жизни, то становится очевидным, что специальные механизмы, сокращающие жизнь, вряд ли могли способствовать успешной борьбе особей за свое существование. Скорее наоборот, те особи, у которых действие подобной программы самоуничтожения было бы нарушено в результате случайной мутации, быстро вытеснили бы всех остальных, поскольку за свою более длительную жизнь они производили бы больше потомства.
Тем более удивительно, что убеждение в существовании специальной генетической программы самоликвидации организма широко распространено и активно пропагандируется. Например, в одном из учебников по биологической кибернетике для студентов изложению подобных представлений посвящен специальный раздел и даже курсивом выделено утверждение об "активном дополнительном регуляторном механизме, который на определенном этапе онтогенеза вступает в борьбу с механизмами обеспечения жизни и прерывает ее течение" [Коган и др.. 1977. с. 65]. Наконец, даже в таком респектабельном издании, как "Британская энциклопедия", можно прочесть следующее: "В коде генетического материала находятся инструкции, которые определяют возраст, свыше которого вид не может прожить даже при наиболее благоприятных условиях" [The New Encyclopaedia Britannica, 1989, vol. 20, p. 471]. Ни одно из этих утверждений не является доказанным, и, более того, как уже отмечалось, подобные утверждения даже теоретически несостоятельны. Однако, учитывая популярность таких представлений, следует более тщательно отнестись к их проверке, не ограничиваясь ссылками на их недоказанность и теоретическую несостоятельность, а обратившись к анализу реальных данных.
На первый взгляд может показаться очевидным, что проверка гипотезы генетически запрограммированной смерти возможна только с помощью прямых молекулярно-генетических, биохимических, цитологических и физиологических экспериментов. Казалось бы, данные о распределении продолжительности жизни организмов в различных популяциях здесь абсолютно бесполезны. Однако при более глубоком анализе проблемы можно предложить, по крайней мере, два разных способа использования таблиц продолжительности жизни, позволяющих провести достаточно надежную проверку гипотезы запрограммированной смерти.
Первый способ проверки основан на сопоставлении таблиц продолжительности жизни особей одного вида, полученных в естественных (диких) и искусственных (лабораторных, домашних, цивилизованных) условиях. Если проверяемая гипотезы верна, то между такими таблицами не должно быть слишком больших различий по срокам жизни взрослых особей. Действительно, чтобы программа самоликвидации могла возникнуть, закрепиться и сохраниться в ходе эволюции, она по меньшей мере должна иметь возможность хоть как-то проявиться в естественных (диких) условиях. Следовательно, возраст включения такой программы должен быть не очень большим, иначе в результате высокой смертности в диких условиях от хищников, голода и инфекций и суровых природных условий просто никто не доживет до рокового возраста и механизм самоликвидации не успеет даже проявиться. Отсюда следует, что продолжительность жизни даже в самых благоприятных условиях не может существенно превышать тех сроков, до которых в диких условиях доживает наиболее выносливая часть популяции, если, конечно, проверяемая концепция верна.
Теперь, если мы обратимся к анализу реальных данных, мы увидим картину, прямо противоположную тому, что следовало ожидать из проверяемой концепции, продолжительность жизни организмов в благоприятных для долголетия условиях намного превышает те сроки жизни, которые наблюдаются в естественных (диких) условиях. Ввиду обилия подобных фактов, ограничимся лишь некоторыми примерами.
Известно, что в благоприятных условиях зяблик (Fringilla coelebs) может прожить 29 лет [Комфорт, 1967]. Однако в диких условиях это практически исключено, поскольку вследствие голода, холода, болезней и нападений хищников около половины птиц погибает в течение одного года, и средняя продолжительность их жизни составляет всего 1.4-1.5 года [Паевский. 1985]. В результате такой высокой смертности до возраста 11 лет доживает лишь 0,1% исходного числа зябликов [Там же].
Аналогичные результаты получены и для полевок (Mircrotus arvalis Pall). В лабораторных условиях средняя продолжительность жизни полевок составляет 7-8 месяцев, причем отдельные экземпляры доживают до 25 месяцев. В диких же условиях средняя продолжительность жизни полевок составляет всего 1,2 месяца, а до возраста 10 месяцев доживает только 0,1% исходного их числа [Фенюк, Шейкина, 1940]. Таким образом, если оценить возраст запрограммированной смерти по продолжительности жизни в лабораторных условиях, то становится очевидным, что никакая программа самоликвидации не могла ни возникнуть, ни сохраниться в эволюции, хотя бы уже потому, что она никак не могла проявиться в естественных условиях, где практически никто не доживает до таких возрастов.
Тот же вывод получается при анализе данных по продолжительности жизни человека. В настоящее время средняя продолжительность жизни в развитых странах достигает 70-80 лет, а подтвержденный рекорд долголетия составляет 120 лет. Если мы примем эти числа в качестве оценки возраста включения программы самоликвидации, то мы должны признать, что подобная программа никак не могла проявиться в эволюции человека, поскольку, согласно данным палеодемографии, до таких возрастов практически никто не доживал. Например, по данным Ачади и Немешкери [Acsadi, Nemeskeri, 1970], возраст, до которого доживала только половина всех родившихся, составлял в позднем палеолите всего 8-9 лет. а в неолите - 26 лет. Более того, даже в эпоху средневековья (IX-XII в.) ожидаемая продолжительность жизни при рождении не превышала 27-29 лет. Изучение скелетов американских индейцев показало, что всего два века назад до возраста 50 лет доживало только 4% населения [Owsley, Bass, 1979]. Для сравнения отметим, что сейчас вероятность дожития до этого возраста в развитых странах составляет 94-96%. Если сопоставить эти факты, то трудно удержаться от вопроса: неужели гарантированное уничтожение нескольких случайно уцелевших и обреченных в диких условиях стариков может быть достаточным эволюционным основанием для формирования и сохранения в геноме человека специальной программы самоликвидации? С этих позиций абсурдность гипотезы запрограммированной смерти становится очевидной.
Приведенные факты, несмотря на их принципиальное значение, могут, однако, показаться недостаточно убедительными для тех, кто скептически относится к теории эволюции и склонен усматривать в ограниченных сроках жизни промысел божий. Поскольку время нашей жизни ничтожно по сравнению с характерными временами биологической эволюции, аргументы, построенные на эволюционных соображениях, для таких людей могут показаться слишком абстрактными и гипотетичными. Нам приходилось сталкиваться и с такой позицией: "Все это было настолько давно, что теперь уже никто не может знать, как оно было на самом деле". И хотя приведенные выше эволюционные аргументы неоднократно высказывались, начиная с 50-х годов нашего века [Medawar, 1957; Williams, 1957], эти аргументы принимаются далеко не всеми исследователями. Поэтому имеет смысл рассмотреть и другой подход к проверке гипотезы запрограммированной смерти, никак не связанный с эволюционно-историческими представлениями.
Второй способ проверки данной гипотезы состоит в изучении характера зависимости смертности от возраста [Гаврилов, 1984а; 19846]. Если проверяемая гипотеза верна, то момент включения программы самоликвидации должен сопровождаться резким скачком смертности. Тщательная проверка подобного предсказания уже была проведена в данной книге (раздел 4.2) и показала, что никакого катастрофического роста смертности в районе предполагаемого предела длительности жизни не происходит. Этот факт опровергает представление о том. что "в коде генетического материала находятся инструкции, которые определяют возраст, свыше которого вид не может прожить даже при наиболее благоприятных условиях" [The New Encyclopaedia Britannica, 1989, vol. 20, p. 471].
Изучение возрастной динамики смертности позволяет проверить и более сложные варианты гипотезы запрограммированной смерти. Так. существует представление, что подобная программа "вступает в борьбу с механизмами обеспечения жизни" сразу после 40-50 лет, но "прерывает ее течение" лишь через 20-40 лет изнурительной борьбы (иначе невозможно согласовать данную гипотезу с тем общеизвестным фактом, что средняя продолжительность жизни людей в развитых странах составляет теперь 70-80 лет). В качестве иллюстрации подобного представления приведем одно характерное высказывание, в котором гипотеза пострепродуктивного саморазрушения выдается за общепризнанный факт: "Общеизвестно, что в жизни человека период старения наступает вслед за периодом расцвета половой деятельности. Именно после 40-50 лет появляются в облике и поведении человека постепенно нарастающие признаки приближающейся старости. Все исследования в области геронтологии приводят к совпадающим результатам, что именно начиная с этого возраста развиваются деструктивные изменения в органах на физико-химическом, клеточном и тканевом уровнях, падает эффективность вегетативных функций, снижаются физическая сила и умственные способности человека. Может быть, эффекты "омоложения", Достигаемые с помощью половых гормонов... цитотоксической сыворотки... недостаточного питания... помимо их общего стимулирующего действия, в какой-то мере связаны и с отсрочкой развития функции саморазрушения" [Коган и др., 1977, с. 67].
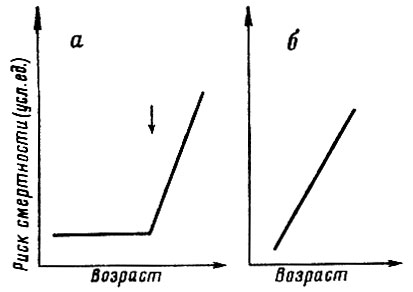
Рис. 31. Два возможных типа возрастной динамики смертности. а - возрастная динамика смертности, ожидаемая в случае справедливости гипотезы активного программированного саморазрушения организма. Стрелкой отмечен момент включения программы самоликвидации; б - возрастная динамика смертности, ожидаемая в случае справедливости гипотезы постепенного накопления дефектов и износа
В случае справедливости подобных утверждений следует ожидать, что в момент включения программы саморазрушения (40-50 лет) должен наблюдаться излом в возрастной динамике смертности, знаменующий собой начало нового этапа ее ускоренного роста (рис. 31, а). Если же никакой программы саморазрушения не существует, а рост смертности с возрастом обусловлен просто накоплением разнообразных дефектов и повреждений (износ в широком смысле этого слова), то следует ожидать монотонного роста смертности с возрастом без каких-либо изломов (рис. 31, б). Иначе говоря, в случае справедливости гипотезы программы в возрасте 40-50 лет должно происходить качественное изменение возрастной динамики смертности, в то время как гипотеза износа допускает возможность расчета смертности в старших возрастах путем простой экстраполяции тенденций смертности, наблюдаемых в младших возрастах (до 40-50 лет).
![Рис. 32. Реально наблюдаемая возрастная динамика смертности. Зависимость интенсивности смертности от возраста женщин Щвеции в 1980 г. [см : Befdkningsforandringar, 1980]](pic/000158.jpg)
Рис. 32. Реально наблюдаемая возрастная динамика смертности. Зависимость интенсивности смертности от возраста женщин Щвеции в 1980 г. [см : Befdkningsforandringar, 1980]
Обратившись к реальным данным (рис. 32), нетрудно заметить, что никакого излома динамики смертности в возрастах 40-50 лет не наблюдается, и более того, смертность в старших возрастах может быть легко оценена путем простой экстраполяции тенденций, наблюдаемых в возрастах 30-40 лет. Само же снижение жизнеспособности, проявляющееся в увеличении риска гибели с возрастом, становится заметным не с 40-50 лет, а гораздо раньше - уже с 10-15 лет. Именно с 10-15-летнего возраста начинается возрастное увеличение общей смертности, смертности от ряда "эндогенных" причин смерти, и появляются первые признаки дегенеративных изменений (например, атрофия тимуса). Впрочем, при более тщательном исследовании не исключено и более раннее выявление неблагоприятных изменений.
Следует отметить, что против гипотезы запрограммированности продолжительности жизни свидетельствует даже сам вид распределения по срокам жизни, описываемый законом Гомперца-Мейкема
Действительно, это распределение хорошо известно в теории надежности под названием первой асимптотической функции распределения наименьших значений (Гумбель, 19651. Как уже отмечалось, это распределение описывает разрушение технических объектов и явления электрического пробоя [Kase, 1953; Гумбель, 19651. Таким образом, динамика смертности организмов совпадает с динамикой разрушения материалов, что свидетельствует в пользу гипотезы износа в самом широком смысле этого слова.
Противники гипотезы износа нередко приводят следующий аргумент. Технические системы, в отличие от систем биологических, по их мнению, не имеют видового предела продолжительности жизни. Этой точки зрения придерживается, в частности, геронтолог А. Комфорт: "Постоянство видового возраста у стареющих форм служит сильным аргументом против примата "механического" старения" [Комфорт, 1967, с. 58]. Однако, как мы уже убедились, никакого видового предела продолжительности жизни, по-видимому, вообще не существует. Поэтому данный аргумент противников гипотезы износа нельзя считать убедительным.
Итак, анализ выживаемости организмов позволяет надежно отбросить гипотезу запрограммированности Продолжительности жизни и сделать вывод о бесперспективности поисков специального "гормона смерти". При этом, однако, следует сделать несколько существенных оговорок. Во-первых, износ одних систем организма может приводить к увеличению нагрузки и ускорению износа других его систем. Таким образом, в организме может возникать каскад зависимых отказов, имитирующий картину его саморазрушения. Во-вторых, при глубоко зашедшем разрушении организма могут включаться механизмы аварийного регулирования, при котором происходит изменение структуры системы с частичным отказом от выполнения основной ее цели [Гублер, 1978]. В этой ситуации аварийные регуляторные реакции по своим проявлениям могут быть внешне очень похожи на процессы саморазрушения организма. И наконец, в-третьих, пока не следует исключать принципиальную возможность существования программы активного саморазрушения организма у некоторых экзотических видов, сложившейся в результате специфики их экологии. Жизнь настолько разнообразна в своих проявлениях, что возможны исключения даже из самых общих закономерностей.
|
ПОИСК:
|
При использовании материалов проекта обязательна установка активной ссылки:
http://gelib.ru/ 'Геронтология и гериатрия'