В. Экспериментальные исследования
Вилочковая железа совершенно правильно рассматривается как железа детского возраста. Довольно часто ставится проблема лечения процессов старения экстрактом вилочковой железы, а также вопрос о роли инволюции этого органа в старении организма.
Принимая во внимание все вышесказанное, вполне ясным становится интерес, уделяемый исследованию влияния экстрактов указанного органа на происходящие в организме химические изменения, в частности, воздействие этих экстрактов на тканевое дыхание*.
* (Исследования были проведены совместно с Н. Апостол.)
В предпринятых нами исследованиях мы применяли гидролизат вилочковой железы, приготовленный Иоаной Милку в отделении эргонологии нашего Института.
Мы подвергли исследованию воздействие указанного экстракта на тканевое дыхание при помощи метода, предложенного Варбургом. Изучалось дыхание мышцы, печени и почки в фосфатной среде, а также в бикарбонатном растворе Кребса, лишенном фосфатов, в присутствии и отсутствии глюкозы.
Мы обнаружили активизацию дыхания вслед за добавлением экстракта вилочковой железы к мышцам и печени как при применении раствора Кребса, так и в фосфатной среде, в присутствии или отсутствии глюкозы.
Что касается почки, то влияние среды видоизменяет ответную реакцию ткани. В среде, лишенной фосфатов, не наблюдается реакции, в то время как в среде с присутствием фосфатов активирующее влияние экстракта вилочковой железы выражено вполне отчетливо. Следует подчеркнуть это влияние среды на ткань почки, деятельность которой тесно связана с качественными и количественными изменениями содержания фосфора.
С целью выяснения того, в какой степени различные невроэндокринные условия могут влиять на это стимулирующее действие экстракта вилочковой железы в отношении потребления кислорода, исследования проводились на супрареналэктомированных, кастрированных, тимэктомированных животных, подвергнутых введению метилтиоурацила. Для уяснения трофической роли нервной системы мы пользовались денервированными мышцами. Принимая во внимание, что коррелятивные отношения между лимфоидными тканями, вилочковой железой и надпочечниками были установлены К. М. Быковым посредством метода условных рефлексов, а также и нами лично, совместно с нашими сотрудниками посредством других методов исследования (морфологических, химических), представляло интерес проследить влияние экстракта вилочковой железы на супрареналэктомированных животных.
Мы установили следующие изменения: в мышщах супрареналэктомированных животных, погруженных в раствор Кребса, стимулирующее воздействие экстракта вилочковой железы сохраняется; в фосфатной среде оно становится непостоянным и статистически недостоверным как в присутствии, так и в отсутствии глюкозы.
В печени, погруженной в раствор Кребса, отмечается реакция, как и у нормального животного. В фосфатной среде, в отсутствии глюкозы, реакция слабо выражена у супрареналэктомированных животных. В присутствии глюкозы реакция не наблюдается.
Все же способность печеночной ткани использовать глюкозу не исчезает в фосфатной среде у супрареналэктомированных животных.
Несмотря на это, присутствие фосфатов и глюкозы в мышще и печени супрареналэктомированных животных препятствует активации потребления кислорода, достигнутной прибавкой экстракта вилочковой железы к рабочему раствору.
В отношении почек различие влияния экстракта вилочковой железы в зависимости от применяемой среды сохраняется и у супрареналэктомированных животных.
Не поддается никакому сомнению наличие тимогенных взаимоотношений. Кастрация изменяет тканевое дыхание.
В наших опытах отмечались статистически достоверные снижения дыхательного коэффициента после кастрации лишь в случае применения раствора Кребса. В фосфатной среде найденные значения находятся в пределах, относящихся к нормальным животным.
У кастрированных животных не отмечается влияния экстракта вилочковой железы на дыхание мышечной ткани ни в растворе Кребса, ни в фосфатной среде.
В печени, в случае применения фосфатной среды, реакция становится недостоверной с статистической точки зрения. Нам не удалось обнаружить никакого активирующего влияния на почки ни в растворе Кребса, ни в фосфатной среде, где реакция происходит с большой интенсивностью у нормальных животных.
У животных с удаленной вилочковой железой мы не отметили никаких серьезных изменений.
У животных, подвергнутых обработке метилтиоурацилом, не наблюдается влияния экстракта вилочковой железы на мышечную ткань в растворе Кребса без глюкозы. В присутствии глюкозы реакция становится, однако, положительной.
К вышесказанному прибавим также то, что при денервировании мышечной ткани отмечается выпадение влияния экстракта вилочковой железы (через 20 дней после операции).
X. С. Коштоянц констатировал, что после денервирования мышцы наблюдается изменение хроматограмм гликогена по отношению к норме.
Экстракт вилочковой железы принимает участие в инсулиновом гликогенезе мышц посредством торможения гликогенетического процесса. Возможно, что структурные изменения гликогена модифицирует влияние этого экстракта.
Аналогичные исследования были проведены нами также с помощью экстракта шишковидной железы. Этот экстракт не оказывает никакого воздействия на дыхание мышечной и печеночной ткани.
На почку экстракт шишковидной железы действовал так же, как и экстракт вилочковой железы.
Не отмечается воздействия на дыхание мышечной и печеночной ткани у супрареналэктомированных животных.
Что касается почек, то как в растворе Кребса, так и в фосфатной среде наблюдается активизация потребления кислорода, как и у нормальных животных.
У кастрированных животных под влиянием экстракта шишковидной железы мы наблюдали незначительную активизацию дыхания мышечной ткани в растворе Кребса. В фосфатной среде реакция не имеет места.
В печени не отмечается особых изменений под воздействием того же экстракта.
В почке реакции отличаются от реакций нормального животного; отмечается отсутствие активирующего влияния как в растворе Кребса, так и в фосфатной среде.
Влияние экстракта шишковидной железы отрицательно также в денервированной мышце.
Из вышесказанного вытекает, что действие экстракта шишковидной железы на тканевое дыхание, при прочих равных условиях, отличается от действия экстракта вилочковой железы в отношении мышечной и печеночной ткани, в то время как в отношении почек оба экстракта оказывают то же воздействие на нормального, супрареналэктомированного и кастрированного животного.
Полученные результаты выдвинули проблему специфичности действия исследуемых экстрактов.
Было сделано несколько попыток для того, чтобы удостовериться, связано ли активирующее влияние дыхания с процессом ферментной активности. С этой целью мы изменили субстрат посредством добавления лактата, пирувата, сукцината, однако полученные результаты оказались отрицательными.
Добавочные исследования показали также, что полученные результаты нельзя объяснить присутствием аминокислот, появляющихся вследствие щелочного гидролиза тканей, так как их количество слишком незначительно. Соображения того же порядка исключают участие гистамина.
По мнению некоторых авторов, дыхание тестикулярной ткани в среде, лишенной глюкозы, снижается одновременно с увеличением возраста животного. Прибавление глюкозы к данной среде вызывает активацию потребления кислорода.
Мы вновь выдвинули эту проблему и одновременно исследовали влияние некоторых воздействий на тканевое дыхание яичек.
Мы исследовали влияние экстракта вилочковой железы, тироксина (в анаболической дозе) и метилтиоурацила, являющегося антагонистом тиреоидной функции.
Изучалось также действие экстракта вилочковой железы на дыхание тестикулярной ткани in vitro как у нормального животного, так и у животных подвергнутых вышеперечисленным воздействиям.
Работы проводились в фосфатной среде в присутствии и в отсутствии глюкозы при рН равном 6,8. Исследовалось также влияние других гексоз.
Подопытные животные (крысы) были разбиты на 3 группы. Первая группа состояла из животных с весом тела от 60 до 100 г, вторая группа - от 100 до 150 г, третья группа - от 150 до 200 г.
Количественные определения проводились посредством манометрического метода Варбурга, а результаты выражались в мм3/мг сухой ткани/час.
Изучались также изменения содержания воды в яичках; было установлено повышение содержания воды, параллельно со старением животного.
Наши исследования позволили нам сделать следующие заключения:
В противоположность утверждениям некоторых авторов, дыхание тестикулярной ткани не изменялось параллельно с возрастом животного в среде, лишенной глюкозы.
Прибавление глюкозы увеличивает дыхание тестикулярной ткани. Указанное повышение менее значительно у молодых животных.
Введение экстракта вилочковой железы приводит к повышению тканевого дыхания яичек молодых животных, при условии отсутствия глюкозы в среде.
Воздействие экстракта вилочковой железы in vitro выражается повышением потребления кислорода как в среде, лишенной глюкозы, так и в глюкозосодержащем растворе.
Тироксин в анаболических дозах тормозит влияние экстракта вилочковой железы в присутствии глюкозы.
Метилтиоурацил, а также тиреоидэктомия снижают возможность тестикулярной ткани потреблять глюкозу.
Изучалось также влияние фруктозы и манозы на дыхание тестикулярной ткани.
Отмечалось повышение дыхания вслед за прибавлением указанных гексоз, как и после добавления глюкозы.
После тиреоидэктомии не отмечается повышения дыхания, даже после прибавления фруктозы или манозы.
Нам удалось установить возможность коррелятивных взаимодействий между вилочковой железой, щитовидной железой и половыми железами, обусловленных их влиянием на углеводный обмен. Мы считаем, что это коррелятивное взаимодействие координируется и регулируется посредством центральной нервной системы.
В рамках экспериментальных исследований Института гериатрии мы изучали также связанные с обменом изменения кожи животных в зависимости от возрастного периода. Мы проследили дыхание кожи у животных от рождения до достижения веса тела, соответствующего взрослому возрастному периоду. Было обнаружено снижение дыхания кожи параллельно с увеличением возраста, а также снижение содержания воды. Было установлено наличие активной сукциноксидазы в коже. Влияние указанного фермента сохраняется в продолжение всей жизни животного. Прибавление солей пировиноградной и молочной кислот не оказало влияния на потребление кислорода, так что нам не удалось установить наличия соответствующих ферментов в коже.
Кастрация крыс-самок приводила к снижению содержания воды и интенсивности дыхания кожи. Удаление шишковидной железы и кастрация также приводят к снижению этих констант, однако значения остаются на более высоком уровне, нежели после кастрации.
Снижения содержания воды и интенсивности дыхания кожи параллельно с возрастом, отмеченные нами, подтверждают данные советских авторов А. В. Нагорного* и Рубановской.
* (А. В. Нагорный, Старение и долголетие, 1950.)
Для уяснения обменного влияния белковых экстрактов вилочковой и шишковидной желез в связи с возрастом животных, мы изучили* действие указанных экстрактов на изменения общих белковых и небелковых азотистых веществ, а также изменения общих белков и воды.
* (Совместно с Н. Апостол.)
Изменения указанных констант были изучены в различных тканях, как например печени, мышцах, сердце, почках, яичках и селезенке при старении животных. Полученные данные позволили нам утверждать, что изменения указанных констант различаются в зависимости от ткани, причем темп изменений характерен для каждой ткани по мере постарения организма.
Лечение экстрактом вилочковой и шишковидной желез приводит к различным результатам.
Наиболее достоверные изменения, наблюдающиеся при воздействии экстракта шишковидной железы, относятся к колебаниям содержания белков в яичках и, таким образом, подтверждают мысль о коррелятивной связи между шишковидной железой и гонадами.
Под влиянием экстракта вилочковой железы происходят изменения в белковом и небелковом обмене яичка, а также в небелковом азотистом обмене селезенки.
Эти результаты были частично пересмотрены в другой работе, где мы изучали изменения небелковых азотистых веществ селезенки и вилочковой железы под влиянием лечения экстрактом вилочковой железы. Была установлена коррелятивная связь между надпочечниками и лимфоидной тканью, селезенкой и вилочковой железой посредством изучения данных корреляций в рамках изменения небелкового азота у нормальных и супрареналэктомированных животных.
При изучении некоторых биохимических данных на белой крысе с удаленной вилочковой железой через 13 дней после рождения мы проследили обмен нуклеиновых кислот, кислоторастворимых фосфорных фракций, общих липидов в печени, селезенке, головном мозгу и мышечной ткани.
Животные были забиты через 60 дней после операции, одновременно с контрольной группой. Из полученных результатов можно вывести интересные заключения относительно углеводного и жирового обмена, а также белкового обмена печени.
В мышечной ткани крысы с вилочковой железой, удаленной вскоре после рождения, неполноценный обмен углеводов, а также нагрузка жировыми веществами выявляют выраженную функциональную недостаточность.
В печени этих животных наблюдается значительное снижение содержания тимонуклеиновой кислоты; это также подтверждает, что вилочковая железа в этом возрастном периоде оказывает первостепенное воздействие на один из конституциональных факторов печеночной клетки.
В головном мозгу и селезенке не отмечалось никаких статистически достоверных изменений.
В нашем Институте изучение шишковидной железы проводится с самых разнообразных точек зрения. Одной из сторон изучения шишковидной железы является ее биохимическое исследование.
Экспериментальные исследования, произведенные нашим биохимическим коллективом*, имели своей целью выяснить влияние шишковидной железы на животных различного возраста, причем полученные результаты выявили соотношение шишковидной железы с процессом роста и другими эндокринными железами, а также влияние этой железы на различные стороны обмена веществ. Полученные результаты показали возможность воздействия на метаболизм в различных отношениях, зависящих от возраста, посредством введения экстракта шишковидной железы, подтвердив биохимическим путем эндокринный характер шишковидной железы.
* (И. Потоп, В. Боеру и сотрудники.)
В первой серии опытов мы изучили обменные изменения, происходившие под воздействием экстракта шишковидной железы у крыс и морских свинок двух различных возрастов (молодые и взрослые), а также у птиц.
Во второй серии опытов на первом этапе мы изучали влияние оперативного удаления шишковидной железы параллельно с введением эпифизгормона молодым, в остальном здоровым животным, а также животным, лишенным коры мозга, оставив для второго этапа задачу исследования тех же констант на животных различного возраста.
Полученные результаты показали следующее: под влиянием экстракта шишковидной железы (вводившегося ежедневно в количестве 1 мл = 0,1 г свежей железы в продолжение 30 дней) происходит активация повышения веса тела, зависящего от возраста и вида животного.
Так, из таблиц 56 и 57, вытекает, что у молодых крыс средняя повышения роста тела равняется 35% у контрольных животных и 50,7% у подопытных животных, в то время как у молодых морских свинок изменение веса тела у животных, подвергавшихся лечению, равняется 25%, а у контрольных достигает 46%. Что касается взрослых животных (морских свинок и крыс), то следует отметить, что повышение веса тела более выражено в контрольной группе, чем в подопытной группе.
Молодые крысы (60 г) с удаленной шишковидной железой прибавили в весе тела за 60 дней после операции меньше чем 25%, в то время как контрольные животные прибавили 126%.
Что касается изменений веса различных органов, то мы отметили следующее: при удалении шишковидной железы у белых крыс наблюдаются вариации веса щитовидной железы и половых желез в положительном смысле, а у леченных эпифизарным гормоном животных - в смысле снижения веса щитовидной железы и половых желез. Эти результаты показали, что шишковидная железа оказывает тормозящее действие на развивающиеся половые железы, что, по-видимому, происходит посредством гонадотропного гормона гипофиза.
Отмечается также активация развития надпочечников, что выражается в повышении веса этих желез у животных с удаленной шишковидной железой и в снижении веса у обработанных животных. В большинстве исследованных случаев отмечалась инволюция вилочковой железы у оперированных животных и увеличение объема желез у обработанных животных. Наличие регрессивного процесса вилочковой железы в результате удаления шишковидной железы говорит в пользу наличия коррелятивных тимоэпифизарных взаимоотношений, вероятно, с участием половых желез. Полученные результаты, подтверждающие данные, ранее зарегистрированные у птиц, говорят, по-видимому, в пользу функционального синергизма между шишковидной и вилочковой железами.
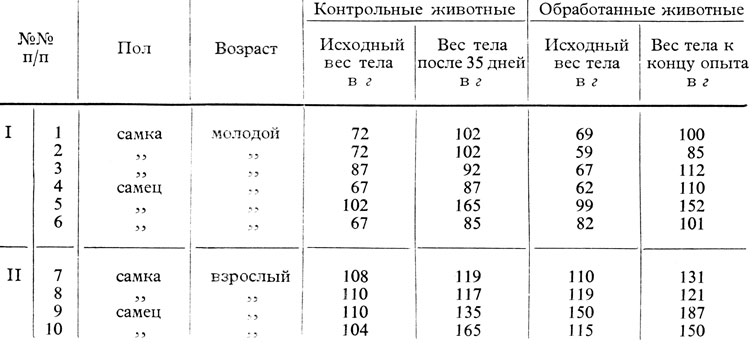
Таблица 56. Крысы
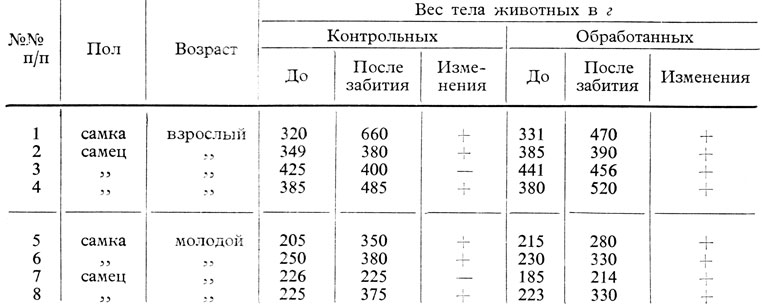
Таблица 57. Морские свинки
Полученные у птиц результаты указывают на сезонные изменения веса щитовидной железы у здоровых птиц. У птиц, которым вводился эпифизарный гормон в зимнее время, отмечалось повышение веса щитовидной железы и снижение веса яичек; в результате введения эпифизарного гормона в весеннее время наблюдалось одновременно снижение веса как щитовидной железы, так и яичек.
Влияние шишковидной железы на обмен веществ было выявлено биохимически посредством изменения содержания минеральных веществ (фосфора, кальция, магния, калия), нуклеиновых кислот, жиров, изменения сахарной кривой на молодой белой крысе с удаленной шишковидной железой и подвергнутой лечению эпифизгормоном (рис. 108 и 109).
Что касается изменений солевого обмена, то при удалении шишковидной железы отмечается повышение содержания фосфора в головном мозгу, печени и вилочковой железе, одновременно со снижением содержания кальция и магния в соответствующих органах; наблюдалось также повышение содержания калия в головном мозгу, одновременно со снижением содержания этого элемента в печени и вилочковой железе. Несколько противоположные результаты отмечались, главным образом, в отношении головного мозга при введении эпифизарного экстракта животным различного возраста.
В крови (у птиц) также наблюдались (таблица 58) различные изменения содержания кальция (повышение), калия, магния и минерального фосфора (снижение) после введения эпифизарного экстракта. Эти данные подтверждают результаты, полученные ранее нашими сотрудниками на птицах.
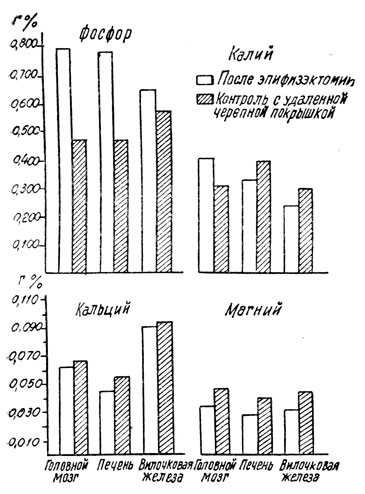
Рис. 108. - Изменение содержания минеральных элементов крови под влиянием удаления шишковидной железы
В наших исследованиях мы показали, что введение эпифизарной или паратиреоидной вытяжки оказывает параллельное влияние на обмен кальция и калия. Встает вопрос, можно ли допустить, что воздействие эпифизарного экстракта происходит посредством околощитовидных желез? Впрочем, эпифизарный экстракт, по-видимому, принимает участие в солевом обмене посредством своего диуретического действия.
В работах, посвященных влиянию возрастного периода на содержание калия в крови и тканях, Г. Бенетато и сотрудники показали, что в старости происходит параллельное снижение функциональной способности и степени растворимости тканевых белков, мобилизация калия из тканей, с его накоплением в крови в виде ионов.
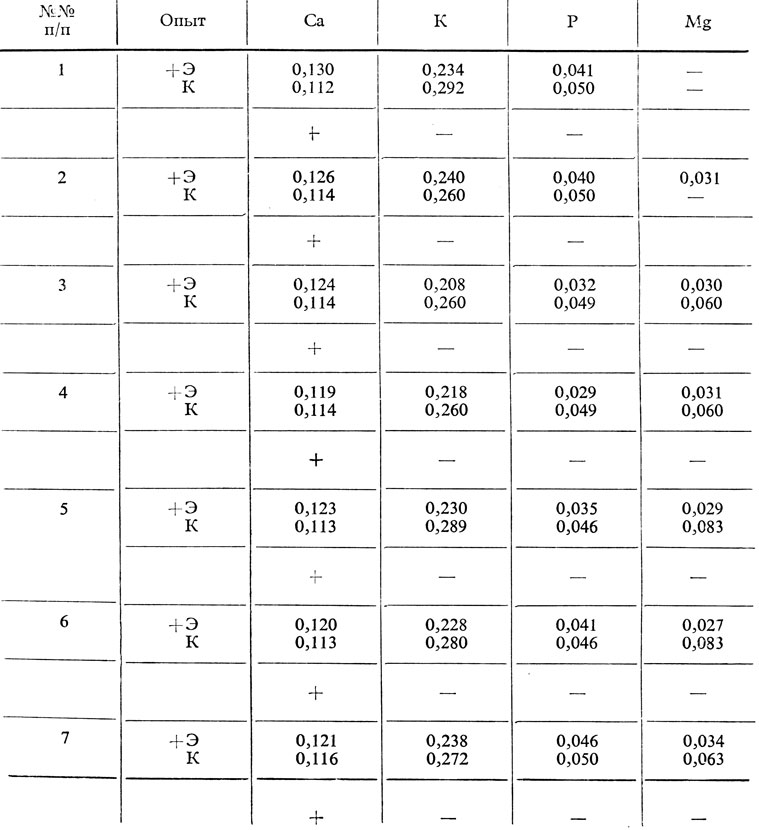
Таблица 58. Содержание в граммах минеральных веществ в сыворотке крови у птиц, обработанных эпифизгормоном (+ Э) и у контрольных животных (К)
В наших исследованиях снижение содержания калия в некоторых тканях - биоэлемента, поддерживающего целостность колоидальных систем клеточной протоплазмы, по-видимому, говорит в пользу идеи экспериментального постарения в результате удаления шишковидной железы. Результаты, полученные посредством изучения различных сторон минерального обмена, подсказывают идею участия шишковидной железы в регуляции указанного обмена веществ, в частности фосфора, калия, магния и кальция.
Исходя из наблюдавшегося нами факта, что при удалении эпифиза происходит активация повышения веса тела и созревания половых желез и торможение указанных процессов при введении эпифизарного экстракта, мы решили проследить изменения содержания нуклеиновых кислот.
Известно, что нуклеиновые кислоты находятся в большом количестве в клетках, синтезирующих значительное количество белков у развивающихся животных, в частности в момент клеточных дифференцировок. Полученные результаты показали изменения, относившиеся к содержанию нуклеиновых кислот у молодых крыс, подвергнутых удалению эпифиза или обработанных эпифизарным гормоном.
Приведенные результаты совпадают с результатами, полученными при изучении морфологических изменений у крыс с удаленной шишковидной железой.
На основании изменения веса этих органов у нормальных животных, со снятой черепной покрышкой, с удаленным эпифизом, подвергнутых обработке эпифизарным экстрактом, можно установить, что при удалении шишковидной железы происходит активация повышения веса яичек, а при введении эпифизгормона в течение 30 дней наблюдается торможение развития этого органа, что выражается в снижений веса яичек. Указанные данные подтверждают результаты, ранее полученные нами совместно с Ш. М. Милку и М. Питиш.
Содержание нуклеопротеидного фосфора изменяется, как показано ниже. При сравнении группы животных с удаленной шишковидной железой с контрольной группой, в которой удалялась черепная покрышка, отмечалось у первых повышение этих веществ на 11,6%, а при сравнении их с нормальными контрольными животными наблюдалось повышение на 25,6%. Обратные результаты отмечались при сопоставлении обработанных животных с нормальными контрольными животными, у которых происходит снижение общего нуклеопротеидного фосфора на 6%.
Результаты этих исследований проводились параллельно с опытами М. Питиш и В. Стэнеску, которые показали, что при удалении шишковидной железы происходит активация сперматогенеза у молодых животных, причем контрольные животные оставались в неполовозрелом возрастном периоде.
Опубликованные в последнее время литературные данные показывают, что синтезу белков предшествует синтез рибонуклеиновой кислоты, которая в свою очередь находится в зависимости от дезоксирибонуклеиновой кислоты клеточных ядер.
Б. В. Кедровский показал, что рибонуклеиновая кислота играет первостепенную роль в клеточных процессах синтеза живой массы, следовательно, во всех процессах развития, дифференциации и деятельности, в частности, в особых функциях некоторых зрелых клеток, среди которых можно упомянуть и о железистой секреции.
Полученные нами результаты подтверждают влияние шишковидной железы на развитие яичек посредством изменения их веса и изменения содержания нуклеопротеидов.
Повышение веса яичек и увеличение содержания нуклеопротеидов при оперативном удалении шишковидной железы, снижение содержания нуклеопротеидов при введении эпифизарного экстракта являются, по-видимому, фактами, указывающими на деятельность пинеальной железы в синтезе нуклеинов на уровне этого органа. С. другой стороны, полученные результаты говорят в пользу наличия коррелятивных связей антагонистического порядка между шишковидной железой и половыми железами.
При исследовании содержания общих липидов в изучаемых нами органах отмечалось, что под влиянием удаления шишковидной железы и введения эпифизарного экстракта наблюдаются противоположные изменения (таблица 59). У животных с удаленной шишковидной железой содержание липидов в головном мозгу, печени и яичках повышается, в то время как при введении эпифизарного экстракта, наоборот, отмечается снижение содержания липидов в указанных органах. Не вдаваясь в подробности, так как число проведенных опытов было слишком незначительным, мы все же можем утверждать, что изменение содержания липидов в исследуемых органах подтверждает влияние шишковидной железы на жировой обмен.
При исследовании гликемии у животных с удаленной шишковидной железой, а также у животных, подвергавшихся лечению эпифизарным экстрактом, отмечаются закономерные изменения: повышение гликемии у крыс с оперативно удаленной шишковидной железой и снижение гликемии у животных, получавших эпифизарный гормон, по сравнению с контрольной группой (рис. 109).
Несмотря на то, что в настоящий момент влияние шишковидной железы на углеводный обмен не подвергается никакому сомнению, следует все же отметить, что механизм ее действия недостаточно выяснен. Возможно, что гипогликемия, вызванная введением эпифизарного экстракта (согласно Ш. М. Милку и сотрудникам), обусловливается резким выделением инсулина, так как хронически вводимый экстракт вызывает гиперплазию лангергансовых островков. Поскольку удаление шишковидной железы влияет на функцию надпочечников, можно предположить, что эта железа играет известную роль посредством обменных гормонов, воздействуя, в частности, на углеводный обмен.
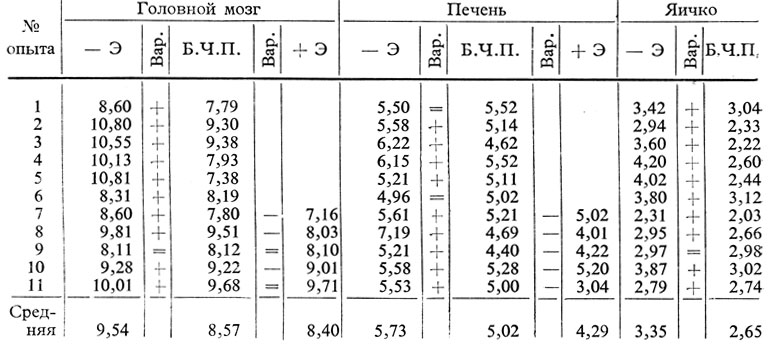
Таблица 59. Содержание общих липидов (в г %) в органах крыс с удаленной шишковидной железой
Равным образом, в связи с возрастной биологией мы изучили экспериментальным путем влияние парентерально введенных нуклеиновых кислот на обменные процессы в органах крыс различных возрастных периодов.
Мы рассматривали изучение влияния рибонуклеиновой и дезоксирибонуклеиновой кислот на обменные процессы как метод исследования, который мог бы выяснить комплексную роль нуклеиновых кислот в целостном организме.
Мы проследили у животных, получавших нуклеиновые кислоты, по сравнению с контрольной группой, различные фосфорные фракции: неорганический фосфор, жирорастворимый фосфор, кислоторастворимый фосфор, а также фосфор, соответствующий нуклеиновым кислотам, и количественные изменения белков; для уяснения механизма действия нуклеиновых кислот мы изучали деполимеразы нуклеиновых кислот, дегидрогеназную активность и дыхание в различных органах подопытных животных.

Рис. 109. - Изменение гликемии у молодой крысы
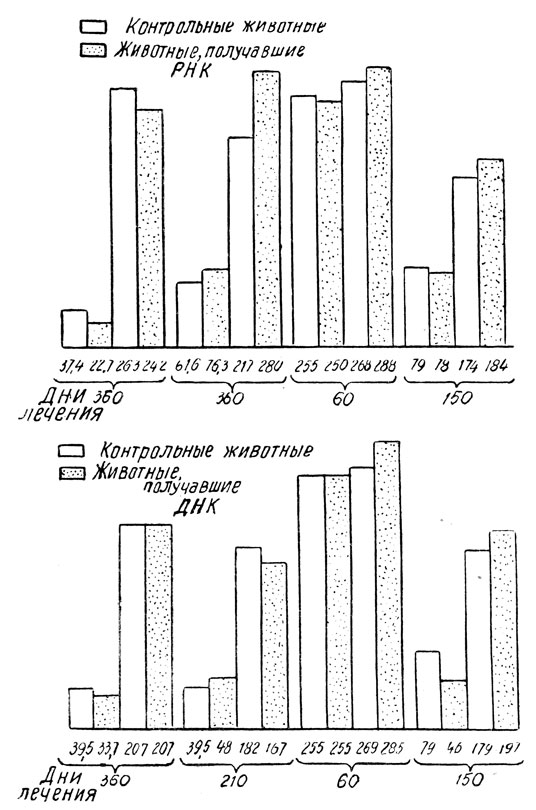
Рис. 110 - Изменение средней веса тела животных, получавших РНК и ДНК
Исследования Глинкова, проведенные под руководством О. Б. Лепешинской, иллюстрируют роль нуклеиновых кислот в обменных процессах и морфогенезе и необходимость изучения воздействия этих кислот in vitro и в живой клетке.
Результаты вышеупомянутых исследований показали следующее:
Животные, которым вводилась рибонуклеиновая кислота, прибавляют в весе по сравнению с контрольной группой. Прибавление в весе происходит в различной степени, в зависимости от возраста животных. Так, у животных, подвергавшихся обработке с 60-дневного возраста, прибавка в весе была большей, чем у старых животных. Животные, получавшие дезоксирибонуклеиновую кислоту, не прибавляли в весе, за исключением животных второго поколения, также старых крыс. Было обнаружено также улучшение общего состояния старых крыс, выражавшееся в прибавке в весе, живости и устойчивости к кожным инфекциям.
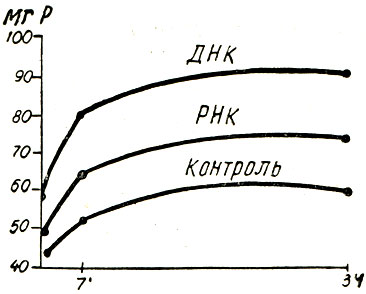
Рис. 111. - Изменение содержания фосфора в головном мозгу при введении РНК и ДНК
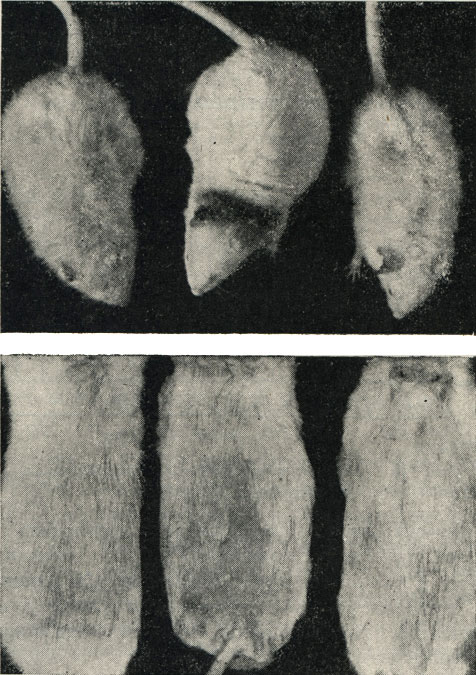
Рис. 112.- Фотография старых крыс: вверху - контрольные животные; внизу - животные, получавшие рибонуклеиновую и дезоксирибонуклеиновую кислоту
Результаты, относящиеся к различным фосфорным фракциям, показывают, что введение рибонуклеиновой кислоты (в продолжение 360 дней) не оказывает влияния на кислоторастворимый фосфор мышечной и печеночной тканей, в то время как дезоксирибонуклеиновая кислота вызывает снижение содержания неорганического фосфора, сопровождающееся повышением общего фосфора, фосфорных эфиров и дифосфоглицериновой кислоты. Дезоксирибонуклеиновая кислота ускоряет, по-видимому, процессы анаэробного распада углеводов в печени. Исследование дегидрогеназной активности печени подтвердило повышение деятельности глюкодегидразы, α- и β-глицерофосфатдегидраз животных, получавших дезоксирибонуклеиновую кислоту. Таким образом, можно предположить, что парентерально введенная дезоксирибонуклеиновая кислота способствует анаэробному распаду углеводов в печени.
В мышечной ткани наблюдается подобный же процесс.
Головной мозг оказался органом с наиболее активной ответной реакцией на действие нуклеиновых кислот.
Как рибонуклеиновая, так и дезоксирибонуклеиновая кислоты вызывают повышение всех фосфорных фракций, за исключением дифосфоглицериновой кислоты (рис. 110 и 111).
Изучение дегидраз показало, что нуклеиновые кислоты повышают дегидрогеназную активность головного мозга. Принимая во внимание, что активность лактикодегидразы и сукциндегидразы были установлены, можно предположить, что нуклеиновые кислоты усиливают в головном мозгу полное превращение угеводов как в анаэробной, так и в аэробной фазах обменного процесса (рис. 112).
|
ПОИСК:
|
При использовании материалов проекта обязательна установка активной ссылки:
http://gelib.ru/ 'Геронтология и гериатрия'