Глава 1. Природа старения и его критерии
I. Измерение степени старения
Старение - это процесс разрушительный. Чтобы измерить степень старения, мы оцениваем снижение жизнеспособности и повышение уязвимости. Возможны и другие способы определения, но в них не принимается во внимание обоснование житейского и научного интереса к процессу старения. Старение проявляется в том, что вероятность смерти с возрастом увеличивается. Изучение старения есть изучение различных для разных групп организмов процессов, которые приводят к повышению уязвимости.
Вероятность того, что отдельный организм, который дожил до времени x, умрет в интервале времени от x до x + 1, зависит от уровня смертности (rate of mortality) (q).
Последний выражается числом индивидуумов, умерших ко времени x + 1, на 1000, доживших до времени x. Сила смертности (force of mortality) (μ) для любого возраста x равна
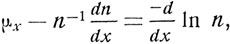
где n - число индивидуумов, доживших до возраста x.
У большинства организмов вероятность умереть в течение определенного отрезка времени подвержена на протяжении жизни колебаниям, часто довольно значительным. Старение проявляется как прогрессивное возрастание (на протяжении всей жизни или после известной стадии) вероятности того, что данный индивидуум умрет в следующую единицу времени под действием случайных причин. Индивидуум больше не в состоянии сопротивляться давлению среды, которому он успешно противостоял в прошлом, хотя само давление и не повысилось. Лишь очень редко мы можем определить уязвимость индивидуума. Наши оценки уязвимости определяются статистически для популяции в целом. Выявление увеличения уязвимости - необходимое условие для выявления
старения. Очевидно, оно будет достаточным только в том случае, если исключить избирательную смертность от внешних причин, связанных с возрастом. В реальных популяциях смерть вызывается как случайными, так и связанными с возрастом причинами. Изменение интенсивности воздействия разных факторов на протяжении жизни человека общеизвестно: взрослый человек подвергается воздействию таких факторов, которые не действуют на детей, и т. п. Различия в степени "риска" на протяжении жизни были изучены и у ряда других животных, таких, как саранча (причины смерти которой изучал Боденгеймер [111]) или галловая муха Urophora [1221]. Пирсон [917] путем разложения кривой выживания для человека на пять составляющих пытался приурочить "старческую смертность" только к одной из этих составляющих, а именно к той, в которой максимум проявляется между 70 и 75 годами. Это решение оказалось бы идеальным, если бы его можно было применять практически. Но метод Пирсона крайне искусствен, и его "пять отдельных смертей" в образе стрелков, направляющих свой огонь на различные возрастные группы, не имеют биологического смысла. В целом, однако, постепенное увеличение силы смертности и уменьшение продолжительности предстоящей жизни для данного возраста в популяции (если исключить различия в интенсивности внешних воздействий) служат показателями старения отдельных ее членов. Предварительный тест для определения старения у данного вида животных основывается, следовательно, на таблицах продолжительности жизни для достаточно больших выборок из популяций, изученных с учетом избирательных причин смерти.
На фиг. 5, А и Б показаны ожидаемые различия в форме кривых выживания для популяций, которым свойственно старение и у которых старение не проявляется. В тех популяциях, где индивидуумы не подвержены старению, а общая смертность носит лишь случайный характер, число особей уменьшается логарифмически, и животные умирают ex hypothesi от причин, которые не зависят от возраста. В популяциях, где индивидуумы умирают только вследствие понижения сопротивляемости, обусловленного старением, кривая состоит из двух ветвей примерно перпендикулярных друг другу (для краткости будет называть ее прямоугольной); после достижения определенного возраста животные умирают под действием таких факторов, от которых они не погибли бы в молодости. В первом случае сила смертности постоянна, а во втором она непрерывно увеличивается с возрастом. Так, у крыс сила смертности после 9-го месяца жизни возрастет в геометрической прогрессии [1269]. Фактически получаемые кривые выживания обычно являются промежуточными между двумя идеальными формами.
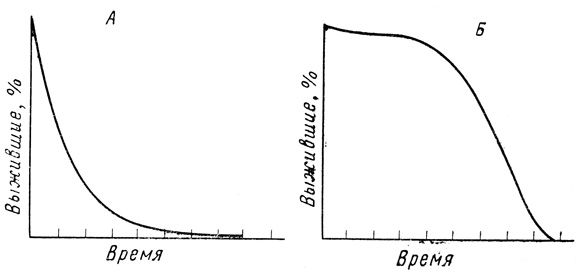
Фиг. 5. Кривые выживания при постоянном уровне смертности (50% в единицу времени) (А) и для популяции, подверженной старению (Б)
Среди наблюдаемых кривых выживания Перл и Майнер [910] различают три основных типа, варьирующих по форме от почти прямоугольных (для организмов с низкой постоянной смертностью на протяжении всей жизни, но умирающих почти одновременно в старческом возрасте) до убывающих логарифмических (для популяций, в которых старение не проявляется или которые вымирают прежде чем оно может появиться) (фиг. 6). Четвертый теоретически возможный тип-тоже прямоугольная кривая, но обратная кривой, характерной для идеальной популяции, которой свойственно старение. В природе он, по-видимому, должен обнаруживаться среди организмов, для которых характерна высокая смертность в раннем возрасте и увеличение продолжительности предстоящей жизни с возрастом. Этот тип кривой выживания характерен для некоторых видов деревьев [1148, 1149], но, вероятно, встречается также и у животных. "Это могут быть животные, у которых продолжительность предстоящей жизни непрерывно увеличивается с возрастом. Так может обстоять дело у многих видов рыб в естественных условиях. Это увеличение продолжается довольно долго. Таким образом, у видов, у которых продолжительность предстоящей жизни равна возрасту или, правильнее сказать, возрасту плюс единица, не могло бы быть вечно живущих особей, но небольшая часть их жила бы очень долго. Столетний человек, знакомый с этими фактами, посочувствовал бы ребенку с продолжительностью предстоящей жизни всего лишь в несколько лет, но позавидовал бы двухсотлетнему" [465].
![Фиг. 6. Типы кривых выживания (Из Пёрла по [14]). А. Тип А: I — голодающая дрозофила (дикая популяция); II — голодающая дрозофила (линия vestigial); III — коловратка. Б. Тип Б: I — таракан, II — дрозофила (дикая популяция); III — автомобиль; IV — мышь. В. Тип В: I — дрозофила (линия vestigial); II — гидра; III — слизень Agriolimax:](pic/000005.jpg)
Фиг. 6. Типы кривых выживания (Из Пёрла по [14]). А. Тип А: I — голодающая дрозофила (дикая популяция); II — голодающая дрозофила (линия vestigial); III — коловратка. Б. Тип Б: I — таракан, II — дрозофила (дикая популяция); III — автомобиль; IV — мышь. В. Тип В: I — дрозофила (линия vestigial); II — гидра; III — слизень Agriolimax:
Старейшая и простейшая попытка выразить математически зависимость смертности от возраста была предпринята Гомпертцом еще в 1825 г.
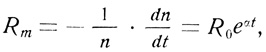
где n - число выживших ко времени t, а a (коэффициент угла наклона) и R0 (гипотетическая смертность в нулевой момент времени) - константы.
Формула Мекхэма отличается от формулы Гомпертца только добавочной константой А, представляющей собой постоянный показатель смертности, независимый от возраста:
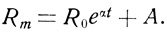
Формула Тейссье описывает кривые выживания, обычные в лабораторной практике, когда в силу малочисленности выборок смертность в течение нескольких первых интервалов времени равна нулю, благодаря чему образуется "плато, характеризующее жизненность взрослых организмов" [1179]:
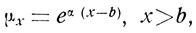
где μx - сила смертности в возрасте x, которая начинает возрастать после того, как пройдено плато протяженностью b.
Ряд более тонких теоретических моделей был предложен как актуариями, которые используют их для прогнозов, так и специалистами по биометрии, пытавшимися сформулировать гипотезу старения. Большая часть их учитывает снижение жизненности, случайное воздействие среды и случайные колебания одного или нескольких физиологических показателей при постоянном или меняющемся гомеостазе. Эти модели подробно рассмотрены Стрелером в двух его обзорах [806, 1138].
![Фиг. 7. Распределение смертей от всех причин по возрастам в экономически развитой стране (А) и смертей пешеходов вследствие несчастных случаев на дорогах (Б) [290, 687]. Зачерненные столбцы - мужчины, заштрихованные столбцы - женщины](pic/000009.jpg)
Фиг. 7. Распределение смертей от всех причин по возрастам в экономически развитой стране (А) и смертей пешеходов вследствие несчастных случаев на дорогах (Б) [290, 687]. Зачерненные столбцы - мужчины, заштрихованные столбцы - женщины
Нарастание уязвимости с возрастом является многосторонним и неспецифическим процессом. Мы можем выразить его в более конкретных терминах. Распределение пешеходов, погибших вследствие дорожных происшествий, по возрастам сходно с распределением по возрастам случаев смерти от всех причин (если исключить ранний детский возраст) (фиг. 7). Этот показатель в значительной степени связан с жизненностью в биологическом смысле, так как он зависит от остроты зрения, слуха и других видов чувствительности, от способности избегать опасности и способности к выздоровлению после травмы.
Но ни один из параметров сам по себе не является достаточным для описания наблюдаемых кривых выживания. Если продолжительность жизни разных видов, например человека и лошади, представить для сравнения на одном рисунке, то наиболее подходящей оценкой, вероятно, будет последний десятый отрезок жизни, так как по нему иногда можно приближенно восстановить ход "физиологической" кривой. Истинным показателем скорости старения, по-видимому, служит производная от силы смертности, поэтому представляется логичным начинать с того, чтобы характеризовать процессы старания показателями смертности или их производными, вычерчивая соответствующие кривые. Но в реальных экспериментах по выживанию все оценки, полученные на основе величины наблюдаемой смертности (qx), дают очень большой разброс, если не брать огромное число случаев, а применение любых стандартных методов выравнивания опирается на предположение о регулярности расположения экспериментальных точек. Небольшое число смертей в раннем возрасте сильно скажется как на ходе кривой выживания (Lx/t), так и на средней продолжительности жизни. В частности, сравнение общей средней продолжительности жизни совершенно непригодно для выражения типа старения, если при этом не сравниваются также причины и распределение смертей по возрастам.
При благоприятных условиях кривые выживания млекопитающих и многих других организмов имеют тенденцию принимать форму, впервые описанную Гомпертцом в 1825 г. Эта "физиологическая" кривая отражает распределение одного из параметров жизненности. Сглаженная кривая выживания, получаемая актуариями, предназначается для предсказания средних показателей. Несглаженные кривые для небольших популяций, хотя они и состоятельны, часто лучше аппроксимируются прямой линией, т. е. приближенно соответствуют уравнению Тейссье [1179]:
где μx - сила смертности в возрасте x, а b<x в этом случае отчетливо выражен период жизненности, характерной для взрослого организма, и кривая начинается с плато, которое соответствует нулевой смертности. Если предположить, что жизненность непрерывно уменьшается в соответствии с некоторым законом (не обязательно линейным), то факторы, постоянно повышающие давление среды или понижающие сопротивляемость по отношению к нему, будут уменьшать b, а факторы, изменяющие продолжительность существования, в распределениях жизненности будут влиять на наклон кривой.
Нири [847] назвал период, предшествующий той точке, в которой в результате падения жизненности индивидуума давление среды становится для него слишком сильным, "индуктивным периодом". Этот период соответствует постоянной, которая введена в уравнение Тейссье [1179].
Если многочисленные факторы, уменьшающие продолжительность жизни, изменяют форму кривой выживания, они обычно уплощают ее сначала до прямой линии, а при еще более сильном действии - до логарифмической кривой. Тогда смертность становится независимой от возраста. Конечный участок имеет тенденцию оставаться на одном уровне, так как последний полудециль погибает только при очень тяжелых обстоятельствах.
Полученное семейство кривых, подобных кривым гистерезиса, образует типичный для животных, имеющих определенную продолжительность жизни, "параллелограмм выживания", по которому можно вычислить точную продолжительность жизни (фиг. 3). Выраженный "хвост", вероятно, возникает вследствие неоднородности анализируемого материала; он может не проявляться при проведении экспериментов на гибридах F1 между инбредными линиями. Кривые этого рода, как обычно считают, отражают увеличение общей смертности, а не изменение скорости старения; по ним нельзя оценить, насколько возрастные изменения, наблюдаемые у выживающих индивидуумов в данный момент, ускорены предшествующими лишениями.
Численность некоторых реальных популяций животных уменьшается приблизительно по логарифмическому закону. "Потенциальное бессмертие" членов популяции, численность которой изменяется подобным образом, - совершенно бессмысленное выражение, вызывавшее в прошлом много волнений у философов, - в практическом отношении значит меньше, чем "потенциальная" встреча двух железнодорожных рельс в бесконечности. Нет ни одной популяции организмов с постоянным общим показателем смертности, которая имела бы в своем составе "потенциально" бессмертных индивидуумов. Единственное преимущество нестареющих организмов по сравнению со стареющими состоит в том, что их шансы умереть остаются постоянными в пределах строго определенного периода, а не уменьшаются с течением времени.
Кривые выживания для 3000 чистокровных кобыл, описанных в Главной племенной книге, очень сходны по форме с соответствующей кривой для цивилизованного человека [234, 235] (фиг. 8, А-В).
![Фиг. 8. Кривые выживания чистокровных кобыл [234, 235, 243]. А. Кобылы, жеребившиеся в 1875-1880 гг. (I и в 1860-1864 гг. (II). Б. Арабские (I) и английские чистокровные (II). В. Сравнение па мастям (данные Главной племенной книги): I - все масти (1860-1864); II - все масти (1875- 1880); III - чалые (200 кобыл)](pic/000010.jpg)
Фиг. 8. Кривые выживания чистокровных кобыл [234, 235, 243]. А. Кобылы, жеребившиеся в 1875-1880 гг. (I и в 1860-1864 гг. (II). Б. Арабские (I) и английские чистокровные (II). В. Сравнение па мастям (данные Главной племенной книги): I - все масти (1860-1864); II - все масти (1875- 1880); III - чалые (200 кобыл)
Эти кривые выживания для человеческого общества с хорошо развитым медицинским обслуживанием и высоким жизненным уровнем имеют форму, промежуточную между прямоугольной и полулогарифмической, но приближающуюся к прямоугольной с начальным спадом, обусловленным детской смертностью. На фиг. 1-3 и 9 приведены, во-первых, кривые выживания для популяций, живущих в нашем веке в различных экономических и климатических условиях, и, во-вторых, изменения в форме кривых выживания для населения северных областей Германии между 1787 и 1800 гг. Много таблиц выживания для популяций, живших до эпохи научной медицины, приведено у Дублина [307]. Значение преимуществ, создаваемых техникой и экономикой, нигде не проявляется так ярко, как при изучении таблиц выживания. Влияние здравоохранения на эти таблицы выражается в большом приближении формы построенных графиков к прямоугольной, а не в удлинении начальной части кривой (до спада). У очень многих организмов, в том числе и у людей, живущих в плохих социальных и медицинских условиях, в раннем возрасте смертность столь велика, что затемняет все последующие изменения, и кривая приближается к кривой четвертого типа по Перлу (перевернутая прямоугольная) (фиг. 6). Увеличение вероятности смерти с возрастом может маскироваться также циклическими изменениями смертности, связанными с размножением и зимовками. Но удостовериться в наличии такого увеличения весьма важно для выявления старения любого организма. Во многих популяциях стареющих животных, например овец и коров (фиг. 14 и 15), кривая выживания взрослых особей не столько прямоугольна, сколько линейна:в течение каждой единицы времени умирает постоянное число индивидуумов, так что смертность неизбежно повышается, поскольку запас животных уменьшается, подобно тому как это было со спутниками Одиссея, постоянное число которых ежедневно поедал Полифем.
![Фиг. 9. Кривые выживания для населения северных областей Германии [1232]. I - данные Хуфеланда [538], полученные путем практических наблюдений и определений; II-III - данные Зильберглейта (II - мужчины, III - женщины), взятые из официальных статистических отчетов, приведенных в Deutsches Statistisches Jahrbuch за 1915 г.](pic/000011.jpg)
Фиг. 9. Кривые выживания для населения северных областей Германии [1232]. I - данные Хуфеланда [538], полученные путем практических наблюдений и определений; II-III - данные Зильберглейта (II - мужчины, III - женщины), взятые из официальных статистических отчетов, приведенных в Deutsches Statistisches Jahrbuch за 1915 г.
Для иллюстрации уменьшения численности в популяции нестареющих животных иногда используют данные о сроках службы фаянсовой и стеклянной посуды (см., например, [793]). Браун и Флуд [147] составили даже таблицу сроков службы стаканов. Для стаканов из прокаленного стекла кривая приближается, правда грубо, к кривой, выведенной для организмов с постоянной смертностью, тогда как уменьшение небольшой группы стаканов из толстого стекла проходило линейно (фиг. 10). Старение, однако, по-видимому, происходит и у стаканов, так как абразия краев облегчает последующее разрушение [147].
![Фиг. 10. Кривые 'выживания' (сроков службы) стаканов из кафетерия [147]. I - стаканы из прокаленного стекла (549 стаканов); II - стаканы из толстого стекла (241 стакан). Одно деление на оси времени соответствует 2 неделям для кривой I и 5 неделям для кривой II](pic/000012.jpg)
Фиг. 10. Кривые 'выживания' (сроков службы) стаканов из кафетерия [147]. I - стаканы из прокаленного стекла (549 стаканов); II - стаканы из толстого стекла (241 стакан). Одно деление на оси времени соответствует 2 неделям для кривой I и 5 неделям для кривой II
Кривые выживания, подобные тем, какие наблюдаются у человека или дрозофилы, удобнее рассматривать как комбинацию, с одной стороны, полулогарифмической "средовой" кривой, обнаруживаемой, когда постоянный показатель смертности высок, а с другой - кривой с прямоугольным падением, обусловленным старением, так как очевидно, что не все индивидуумы, умершие на середине жизненного пути, в какой-то мере обязаны своим несчастьем старческому увеличению уязвимости, как бы рано оно ни началось.
Боденгеймер [111] проводит очень удобное различие между "физиологической" долговечностью вида, которая достигается в оптимальных условиях, в генетически однородной популяции и приближается к наибольшей зарегистрированной у данного вида продолжительности жизни, и "экологической" долговечностью, которая представляет собой среднюю величину, наблюдаемую эмпирически в данных условиях. Идеальная прямоугольная "физиологическая" кривая, постулированная Боденгеймером, - это в лучшем случае удобная абстракция, так как определяющие ее генетические экологические факторы в реальной популяции практически нельзя выявить. Но у некоторых форм в лабораторных культурах или в условиях одомашнивания наблюдаемая кривая выживания приближается к идеальной прямоугольной, причем это приближение особенно сильно выражено в некоторых человеческих популяциях. Однако с позиций статистического понимания старения у большинства лабораторных животных бессмысленно противопоставлять "физиологическое" старенио "патологическому". Если старение рассматривать как повышение общей уязвимости, боденгеймеровская "физиологическая" долговечность занимает на кривой область, в которой повышение уязвимости по отношению ко всем вредным факторам среды происходит настолько резко, что даже серьезная защита против таких факторов не может очень значительно продлить жизнь. Тип кривой может быть изменен, а наблюдаемая физиологическая долговечность увеличена путем устранения тех или иных частных причин смерти, например энтерита или отита у старых крыс [625], но отсрочка смерти, достигаемая таким путем, ограничена сама по себе. Таким образом, рассуждения по поводу существования у млекопитающих "естественной" смерти, не связанной с патологическими процессами, совершенно праздны.
На основании таблиц выживания выявить старение можно лишь при условии достаточно низкой смертности в раннем и зрелом возрасте, благодаря чему число животных, достигающих старости, оказывается достаточно большим; при этом эндогенное повышение чувствительности к случайным факторам, вызывающим смерть, становится очевидным. Так, в естественных условиях смертность мышей настолько высока, что они не доживают до старости, однако в лабораторных условиях кривая их выживания сходна с кривой выживания для населения стран Западной Европы за 1900 г. [465, 706] (фиг. 52). Даже самый тщательный уход не обеспечивает популяции лабораторных животных такие условия, какие создает медицинское обслуживание для современного человека, но, если бы удалось создать такие условия, кривая выживания для этих животных была бы сходна с кривой выживания для населения стран Западной Европы за 1953 г.
Организмы, которые, судя по кривым выживания, подвержены старению, характеризуются видовым возрастом, т. е. средним возрастом в момент смерти, который является характерной чертой вида, если он живет в условиях, приближающихся к боденгеймеровским "физиологическим" условиям.
Медавар [793] недавно вновь указал на ряд недостатков, свойственных статистическому определению старения. Очевидно, что с помощью преднамеренного или даже непреднамеренного отбора материала можно получить практически любую кривую выживания. Весьма критически следует относиться к использованию в качестве показателя непрерывности процесса понижения жизнеспособности данных, учитывающих только возраст в момент смерти. На форму такой кривой влияют многие обстоятельства, в том числе и генетическая гомогенность выборки. Сама частота различных видов "риска" варьирует в разных возрастных группах; так, статистическую картину старения может симулировать, например, кривая выживания для любой популяции рыб в условиях лова их сетью с определенной величиной ячеек. В природе к такому же результату приводит избирательная гибель от хищников. Увеличение силы смертности мужчин призывного возраста во время войны уж никак нельзя считать проявлением старения. Вместе с тем кривая, описывающая частоту некоторых причин смерти, таких, например, как рак [1019], идет параллельно общей кривой смертности. При использовании силы смертности как показателя старения необходимо, как мы видели выше, исключить по возможности внешние факторы, влияние которых с возрастом перестает быть случайным. Однако всего этого нельзя сделать с достаточно строгой логической последовательностью. Что касается кривых выживания для человека, то здесь возможны серьезные изменения причин смерти, приходящиеся на период индивидуальной жизни; конституциональные же различия между индивидуумами в скорости старения гарантируют нам, что генетическая конституция лиц, доживших, например, до 60 лет, не может служить показателем для всей изучаемой группы. В действительности ошибок такого рода можно избежать или же их можно скорректировать для практических целей. Однако их всегда следует учитывать, когда делают выводы относительно старения на основании какой-либо кривой выживания.
Поскольку нет иного прямого способа измерения подверженности индивидуума смерти, кроме фактического его умерщвления, статистическое определение старения, хотя и отражает реальный процесс, происходящий в индивидуумах, можно проверить только на популяции. Для этой цели биолог использует методы, первоначально созданные и усовершенствованные актуариями.
![Фиг. 11. Соотношение между индивидуальной (сплошные кривые) и средней (пунктирные кривые) скоростью роста в период полового созревания [1173]. А. Изменение скорости роста с возрастом у 5 детей. Б. Те же кривые совмещены по максимальной скорости; по шкале абсцисс отложены годы до и после достижения максимальной скорости](pic/000013.jpg)
Фиг. 11. Соотношение между индивидуальной (сплошные кривые) и средней (пунктирные кривые) скоростью роста в период полового созревания [1173]. А. Изменение скорости роста с возрастом у 5 детей. Б. Те же кривые совмещены по максимальной скорости; по шкале абсцисс отложены годы до и после достижения максимальной скорости
В основе различий между статистическим и биологическим подходом к проблеме определения смертности лежат различия между перспективным и ретроспективным использованием статистики. Из этих различий практически самое важное для нас состоит в характере отношения к выравниванию. Почти для всех перспективных целей удобно в первом приближении принять некоторую степень единообразия, чтобы по возможности различить малые и большие модальности, а минорные модальности сгладить. Для большинства ретроспективных целей основная задача заключается в том, чтобы из весьма скудных данных выделить столько надежной или даже суггестивной информации, сколько позволит уровень достоверности. Для этого необходимо применять методы, которые позволяют выявить тонкую структуру там, где она представлена. Основное требование, предъявляемое к биологу, использующему стандартную методику актуариев для исследования возраста, состоит в том, чтобы он был способен четко отличать закономерности, присущие, по-видимому, изучаемому материалу, от тех, которые обусловлены применяемыми методами. Большинство популяций животных слишком гетерогенны для построения кривых выживания. Кривые, полученные на таком материале, являются усредненными. Они пригодны для предсказания пределов, но часто вводят в заблуждение, если их используют в качестве показателей характера биологических процессов. На фиг. 11 показан подобный эффект на кривых роста человека. Здесь изображены кривые индивидуальных скоростей роста как функций возраста у пяти детей; если построить выровненную среднюю для этих кривых, комбинируя измерения так, как это делают 4 А. Комфорт при исследовании популяции методом поперечного сечения, то получается сглаженная кривая (пунктирная кривая на фиг. 11, А), совершенно не отражающая истинного характера индивидуальных кривых скорости роста, имеющих типичный пик. Многие примеры, приводимые для иллюстрации положения, что "природа не делает скачков", получены именно в результате такой обработки. Когда изучение популяции проводят одновременно методом "продольного" и "поперечного" сечения, ошибка становится очевидной. Что касается возрастных изменений жизненности, то провести подобное их изучение мы пока не можем.
Ясно, что в прикладной биологии и особенно в медицине желательно иметь возможность не только делать выводы относительно существования старения у данного вида, но и судить о степени старческих изменений у отдельных индивидуумов. Такая оценка должна основываться на вторичных критериях и может быть сделана точно только для тех форм, как, например, человек или дрозофила, жизненный цикл которых был объектом интенсивного изучения. Значение статистического определения состоит в том, что оно заставляет нас обратиться к исследованию соответствующих популяций. До сих пор нередко поступали так: содержали единственную особь - птицу или крупную лягушку, в течение 10-20 лет, и когда ее находили мертвой, описывали гистологические изменения в ее тканях в статье под названием: "Старческие изменения в нервной системе воробья (или жабы)". Точно так же как нельзя судить о старении по любой кривой выживания, согласно которой сила смертности возрастает, нельзя описывать "старческие" изменения на основании изучения отдельных организмов, принадлежащих к группе, жизненный цикл которой (в смысле старения) совершенно не изучен.
Для организмов, жизненный цикл которых хорошо изучен, кроме кривой выживания, можно использовать в качестве вторичного показателя старения другие критерии; речь идет не о простых измерениях хронологического возраста, основанных на строении чешуек, зубов или отолитов. Некоторые факторы, которые действительно служат прямой мерой жизненности или уязвимости, такие, как смертность от ожогов [57] или число дней нетрудоспособности в течение года [1046], соответствуют общей силе смертности. Предположение об уменьшении скорости заживления ран с возрастом (дю Нойи [867] рекомендовал использовать это уменьшение в качестве меры старения) было основано на исследованиях, проведенных на малоподходящем клиническом материале. Это предположение не было подтверждено позднейшими работами [128, 402], хотя показано, что скорость заживления сквозных ран зависит от возраста [101]. Менее общие критерии, такие, как эластичность кожи у человека [322, 603], вес органов и относительная их гипоплазия у крыс [622, 625], частота сокращений сердца у ветвистоусых раков [377, 378, 557, 795, 796], удой молока у коров [145], яйценоскость у кур [208] (фиг. 19), многие гистологические признаки и интенсивность общего обмена веществ или метаболизма тех или иных соединений имеют значение в строго определенных границах, но все они характеризуются выраженной изменчивостью, не зависящей от общего процесса старения. Например, если, вызвав у ветвистоусых раков задержку в развитии, искусственно увеличить среднюю продолжительность их жизни, то частота сокращений сердца перед смертью не уменьшится до уровня, обычного в старческом возрасте [557]. Майнот [810], Гертвиг и другие считали постепенное уменьшение ядерно-плазменного отношения общим признаком старения организмов; позднейшие работы не подтвердили этого положения, и сегодня его необходимо выразить на более точном биохимическом языке. Дегидратацию прежде также считали общим признаком. Однако, проанализировав старые и вновь полученные данные, Лаури и Гастингс [727] пришли к выводу, что для старчески измененных тканей млекопитающих более характерно повышение степени гидратации, обусловленное, возможно, внеклеточным отеком, разрушением клеток и даже более серьезными патологическими изменениями, например сердечной недостаточностью. В настоящее время нет биологического критерия, характеризующего "старость" тканей и клеток, и поиски такого критерия отражают неправильную в своей основе концепцию. Она состоит в сравнительной оценке всех подобных критериев по отношению к главному проявлению старения, для которого весьма существен статистический подход, - уменьшению устойчивости к случайным факторам, вызывающим стресс. Все утверждения относительно старения, основанные на патологической анатомии или на общих теориях, которые рассматривают его как единый процесс, выглядят довольно сомнительными.
Падение скорости роста на протяжении жизни в некоторых или во всех тканях представляется почти всеобщим свойством многоклеточных.
"Видовая скорость роста всегда падает; живая ткань постепенно теряет способность к самовоспроизведению с той скоростью, с какой она была образована. Майнот сделал это обобщение, которое по праву известно как "закон Майнота", сравнивая полученные им кривые скорости роста, выраженной в процентах. И именно он первым осознал, что точка перегиба на интегральной кривой роста и создаваемое тем самым деление на период положительного и отрицательного ускорения не имеет практического значения. Постепенное рассеяние "энергии роста", утверждаемое этим первым законом, должно, по мысли Майнота, быть выражением старения - "старости" с ее повседневными проявлениями. Старение с этой точки зрения не есть процесс, наступающий после окончания предшествующего периода созревания: старение - это развитие, рассматриваемое с другого конца жизни" (Медавар [792]).
Использование этого критерия, который легко поддается измерению и может быть с соответствующими предосторожностями применен к небольшим группам животных, а также к отдельным тканям и органам, предполагает принятие майнотовского определения старения. Это определение обоснованно. В то же время снижение видовой скорости роста не служит мерой старения в его статистическом смысле, так как оно не параллельно увеличению силы смертности, и даже не является обязательным предвестником старения, если у тех животных, у которых способность к обновлению или росту ряда или всех тканей падает до нуля, смертность увеличивается с возрастом. Речь идет о статистическом, разрушающем старении, и если бы старение у человека сводилось лишь к уменьшению скорости роста, то оно вряд ли было бы предметом всеобщего внимания.
Возможно, что будущие исследования приведут к созданию рабочего и вполне законного метода "прямого измерения" старения индивидуумов, основанного на задержке деления клеток в эксплантатах тканей от старых животных [221, 790, 1145]. До сих пор не выработан практический тест такого рода. Между тем точные наблюдения старения у разных позвоночных почти все проведены на организмах, кривые выживания которых совершенно достоверны (большие рыбы, крокодилы, черепахи). У этих форм относительно легко можно определить (гистологически или путем скрещивания) способность к размножению особей разного возраста. Следовательно, выводы, основанные на старении в отношении способности к размножению, получить сравнительно легко, особенно если вспомнить о непреодолимых трудностях, связанных с измерением силы смертности в таких случаях.
Снижение способности к размножению - весьма общее явление для тех позвоночных, которые подвержены старению, оцениваемому по увеличению силы смертности; к показателям такого снижения у различных форм относятся изменения в половых железах, потеря вторичных половых признаков, прекращение овариального цикла, уменьшение сперматогенеза, понижение способности к оплодотворению, вылупляемости, а также уменьшение числа потомков в отдельных пометах и их жизнеспособности. Однако шкала времени для этих процессов отличается от шкалы увеличения силы смертности, и у разных видов их соотношение различно. Половые железы часто ведут себя как обособленный "организм" со своей собственной детерминированной продолжительностью жизни, но это справедливо равным образом и для других образований, таких, как зобная железа. Ограниченная продолжительность жизни половых желез представляется особой категорией только потому, что по понятиям эволюционной телеологии половые железы - важнейшая часть организма. Старение организма в целом, происходящее после окончания периода размножения, по крайней мере для большинства видов, является процессом, реализуемым только благодаря вмешательству человека, и не рассматривается эволюционной телеологией. Можно спорить с мнением о том, что если у данного вида установлено старение половых желез, то соматическое старение, как правило, неизбежно, так как в пострепродуктивном периоде давление отбора, обеспечивающее гомеостаз, не действует. Прямая физиологическая связь между активностью половых желез, с одной стороны, и ростом и жизненностью животного - с другой, была выявлена у нескольких форм (например, дафния [317, 318]), хотя даже у дафнии овогенез продолжается до самой смерти [1053]. Однако у многих позвоночных даже полная кастрация очень мало или вовсе не уменьшает продолжительность жизни, а иногда даже увеличивает ее. В эволюционном смысле старение половых желез - наиболее важная форма старения; из этого, однако, вовсе не следует, что старение половых желез должно сопровождаться соматическим старением, если только оба эти процесса не связаны друг с другом причинно. Исследование функции размножения ввиду легкости ее измерения остается наиболее достоверным тестом при оценке сохранения жизненности у старых животных; сохранение способности к размножению свидетельствует об отсутствии необратимых изменений в органах одной из важных систем.
Снижение интенсивности метаболизма, либо определяемое непосредственно (калориметрически или манометрически), либо обнаруживаемое по снижению спонтанной активности, также рассматривали как показатель старения; часто на основании теоретических соображений считали, что это снижение отражает накопление неактивной "метаплазмы" взамен активной протоплазмы [589] или завершение "мономолекулярной аутокаталитической реакции", как это постулируют Робертсон [989] и Берталанфи [1306]. Об уменьшении частоты сердечных сокращений у ветвистоусых раков [557] уже говорилось выше. У человека средняя частота сердечных сокращений в покое также уменьшается на протяжении как внутриутробного развития, так и всей дальнейшей жизни. Чайльд [197] измерял возраст гидромедуз по уменьшению у них частоты пульсации. По литературным сведениям у ряда беспозвоночных (планарии [196], гидромедузы [197], моллюски [527, 528]) и в некоторых изолированных тканях позвоночных (хрящи суставов [1004-1006], кровеносные сосуды крысы [693, 694], мышцы птицы [404], мышца кролика [189], гомогенат головного мозга крысы [971], гомогенат тканей печени, почек и сердца [904], лимфатическая ткань мыши [1229]) потребление О2 с возрастом снижается.
Как обнаружили Рокштейн и Гутфройнд, в мышцах комнатной мухи происходит уменьшение содержания аденозинмонофосфата и накопление аденозинтрифосфата, что, по-видимому, обусловлено потерей фосфатазной активности. В калориметрических опытах, проводимых на целых млекопитающих, установлено общее снижение теплопродукции с возрастом [80, 81, 121, 609, 754, 1078, 1079, 1107]. Однако подобно уменьшению энергии роста максимальное уменьшение содержания АТФ происходит в начале жизни; у человека после 50 лет оно идет относительно медленно [1080]. Этот процесс не параллелен увеличению смертности с возрастом. Кроме того, обнаруживаются значительные индивидуальные различия. Кунде и Норлунд [652] не нашли значимого снижения интенсивности основного обмена у собак до 12 лет. У крыс Бенедикт и Шерман [82] при проведении исследований на одних и тех же особях наблюдали небольшое уменьшение теплопродукции с возрастом; однако, поскольку с наступлением старости уменьшается и вес тела, интенсивность обмена на единицу веса, по-видимому, даже увеличивается. Потребление О2 на 1 л межклеточной жидкости у человека не снижается с возрастом [1081]. Более полную библиографию по этому вопросу приводит Шок [1080]. Таким образом, для большинства организмов до сих пор нельзя дать рациональную оценку индивидуального старения на основании изменения интенсивности обмена веществ.
|
ПОИСК:
|
При использовании материалов проекта обязательна установка активной ссылки:
http://gelib.ru/ 'Геронтология и гериатрия'