3. Позвоночные
Возможность вызвать продолжительную, но обратимую задержку роста и развития у млекопитающих посредством недостаточного питания впервые была показана в работах Осборна и Менделя [883, 884]. Хорошо известны исследования Мак-Кея на крысах; он получил непосредственно на млекопитающих данные о таком же влиянии недостаточного питания на рост, какое раньше было установлено для членистоногих. Эти исследования представляют собой единственную удачную попытку разрешить проблему видового возраста у млекопитающих. Указанная проблема является ключевой для медицинской геронтологии, и тот факт, что характер роста у крыс служит скорее исключением для млекопитающих, ни в коей мере не умаляет его значения. Эксперименты, впервые описанные в 1934 г. [738, 739, 741, 1039], в дальнейшем продолжались в течение ряда лет и полностью изложены в обзоре Мак-Кея [737]. Группы крыс содержались на полноценной по своему составу, но недостаточной по калорийности диете, и поэтому их рост замедлялся. После периода задержки, длившегося 1000 дней, калорийность пищи повышалась для возобновления роста. После этого животные быстро вырастали до размеров взрослой особи, несмотря на то что продолжительность их жизни уже превышала норму, характерную для этой линии; животные жили приблизительно в два раза дольше, чем наиболее длительно живущие особи из контрольной группы (без задержки роста) (фиг. 64). Такое увеличение продолжительности жизни сочеталось с уменьшением частоты многих хронических заболеваний, что, по-видимому, отражает действительное уменьшение смертности в старческом возрасте от случайных причин. Это уменьшение происходило главным образом за счет снижения смертности от заболеваний легких и от опухолей. "В общем при задержке роста крысы сохраняют активность и выглядят молодыми, несмотря на значительный возраст. Они очень проворны. На 2-й и 3-й год жизни может возникнуть слепота. Частота пульса у этих животных составляет 340 ударов в 1 мин, т. е. на 100 ударов ниже нормальной" [737]. По интенсивности основного обмена крысы с задержанным ростом занимают промежуточное положение между нормальными молодыми и нормальными старыми животными [531]. У крыс с задержкой роста на 850 дней теплопродукция на единицу поверхности тела была ниже, а на единицу веса выше, чем у контрольных животных [1276]; в аорте и почках у них наблюдалось в общем более выраженное отложение солей кальция, чем у контрольных животных [540], возможно, вследствие относительно более высокого содержания минеральных веществ в их диете [69]. Последующие серии экспериментов с недокормом животных, которые уже достигли зрелости, к сожалению, были усложнены введением многих переменных (моцион, потребление казеина, добавка в диету печени и т. д.). В этих экспериментах недокорм вызывал значительное увеличение продолжительности жизни по сравнению с контрольными животными, получавшими достаточное питание, но различия были гораздо менее выражены, чем в опытах с задержкой роста у молодых крыс. При этом наиболее важными факторами в определении продолжительности жизни были те, которые обусловливали степень ожирения [740, 1084]. Указанное различие объяснялось главным образом повышенной частотой заболеваний почек при высоком содержании белка в пище и при ожирении [1040]. В противоположность результатам, полученным на животных, рост которых был задержан в молодости, у взрослых крыс при недокорме не снизилась частота случаев хронической пневмонии и опухолей [743, 1039].
![Фиг. 64. Кривые выживания нормальных крыс и крыс с задержанным ростом (самцов и самок), показывающие влияние ограничений в питании [743]. I - нормальные крысы (данные 1934 г.); II - нормальные крысы (данные 1943 г.); III - крысы с задержанным ростом (данные 1943 г.)](pic/000077.jpg)
Фиг. 64. Кривые выживания нормальных крыс и крыс с задержанным ростом (самцов и самок), показывающие влияние ограничений в питании [743]. I - нормальные крысы (данные 1934 г.); II - нормальные крысы (данные 1943 г.); III - крысы с задержанным ростом (данные 1943 г.)
По характеру основного обмена крысы с ограниченным питанием занимают промежуточное положение между нормальными молодыми и нормальными взрослыми крысами [1276]. Ограничение их размеров есть не что иное, как ограничение числа клеток: животные с задержанным ростом имеют такую же популяцию клеток, как животные соответствующего размера, а не возраста [384]. Влияние недокорма осуществляется, по-видимому, через посредство гипофиза - пищевая гипофизэктомия [1034] - и устраняется введением гормона роста [534]. Для тканей крыс с задержанным ростом характерен более короткий латентный период в культуре [525], а тепловое сокращение коллагена из хвоста этих животных свидетельствует о том, что его возрастные изменения выражены в меньшей стецени [204]. У нормальных взрослых животных влияние недокорма менее заметно [741].
Росс [1007] изменял содержание каждого компонента в рационе крыс по отдельности. Ограничение в потреблении одних лишь белков вызывало небольшое увеличение продолжительности жизни. Ограничение в потреблении углеводов давало некоторое увеличение продолжительности жизни, но меньшее, чем наблюдалось при ограниченном потреблении белков и углеводов и снижении общей калорийности пищи. Если содержание белков в пище понижено, а углеводов - повышено, крысы сами сокращают потребление такой пищи и живут соответственно дольше, чем животные, не ограничиваемые в пище (фиг. 65).
Крысы с задержанным ростом по химическому составу печени напоминают молодых животных: количество фосфора РНК, извлекаемое реактивом Эдсолла, у 12-месячных особей даже выше, чем у нормальных 1-месячных животных, и обмен белка как будто повышается по сравнению с нормой. При восстановлении нормального питания эти показатели быстро "догоняют" свой хронологический возраст [858].
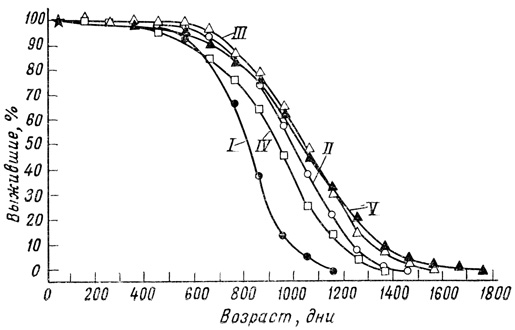
Фиг. 65. Кривые выживания для четырех групп крыс, содержавшихся на ограниченной полусинтетической диете, и для одной группы крыс, получавших вволю продажную диету. I - продажная лабораторная диета; II - диета с высоким содержанием казеина и высоким содержанием сахарозы III - диета с высоким содержанием казеина и низким содержанием сахарозы; IV - диета с низким содержанием казеина и высоким содержанием сахарозы, V - диета с низким содержанием казеина и низким содержанием сахапозы
Результаты этих экспериментов показывают, что по крайней мере некоторые млекопитающие способны переносить продолжительную задержку роста в неполовозрелом состоянии без какого-либо ускорения старения. Задержка эта неполная, так как при полном отсутствии прибавки в весе наступает смерть. Наиболее важный вывод из этих экспериментов, по-видимому, состоит в том, что само по себе старение есть прямое следствие не столько прекращения роста, сколько достижения определенной стадии развития, время наступления которой отчасти, но не полностью связано со скоростью роста ибо нет данных, что голодающие крысы могут оставаться "молодыми" неопределенно долго. Более того, после 1150 дней голодания только половина животных с задержкой роста оказалась способной возобновить рост [743]. В более поздних работах получены данные, что увеличение продолжительности жизни, хотя и в меньшей степени, можно вызвать посредством периодических ограничений в пище, причем это не сопровождается каким-либо явным изменением скорости роста [176]. Более того, большинство изменений, в конечном счете определяющих видовой возраст, появляется в период зрелости, так как у взрослых крыс продолжительность жизни при недокорме увеличивается гораздо меньше [740].
В противоположность данным Мак-Кея задержка роста у крыс, вызванная до прекращения вскармливания материнским молоком, неизменно имеет следствием карликовые размеры и неблагоприятно влияет на продолжительность жизни [1267].
При менее сильном или периодическом недокорме продолжительность жизни увеличивается, но главным образом за счет сокращения ранней смертности; видовой же возраст изменяется в меньшей степени. Ризен и др. [981] нашли, что у крыс линии Вистар долголетие, грубо говоря, пропорционально степени недокорма: голодание в течение 1 дня из каждых 3 или 4 приводит к значительному увеличению средней продолжительности жизни. Выигрыш от задержки роста у крыс при содержании их на смешанной диете проявляется сильнее, чем на вегетарианской [177, 178]. При голодании в течение 1 дня из каждых 3 продолжительность жизни увеличивается в среднем на 20% у самцов и на 15% у самок без остановки роста [176]. Томассон установил, что кривая выживания у крыс, получающих с пищей 27% жира в виде коровьего масла, шла наклонно с небольшим начальным плато. После замены животного жира растительным (рапсовое) плато удлинилось примерно на 25 недель и кривая по форме приближалась к прямоугольной; рост замедлялся главным образом из-за плохого аппетита. Кроме того, заметно снижалась частота заболеваний почек. В работе Берга и Симмса [86] при умеренном сокращении рациона до уровня, при котором рост не подавлялся и половое созревание не запаздывало, продолжительность жизни крыс увеличивалась на 25% (200 дней) у самок и на 30% (300 дней) у самцов с соответствующим более поздним появлением заболеваний и опухолей.
Эксперименты такого же рода были проведены на мышах и дали очень сходные результаты. Как общую продолжительность жизни, так и продолжительность репродуктивного периода можно увеличить путем снижения калорийности пищи. При стандартной мышиной диете, содержащей половину нормы калорий, самки линии C3H, которые обычно становятся стерильными уже в 11-12 месяцев, давали пометы еще в возрасте 21 месяца. Рекорд принадлежит мышам, которых ограничивали в пище в течение 11 -15 месяцев, а затем перевели на полноценный рацион [179]; голодание в течение 2 дней в неделю с добавлением или без добавления в пищу нуклеиновых кислот приводило у альбиносов к увеличению продолжительности жизни на 50-60% [991]. Опыты на мышах особенно удобны благодаря доступности инбредных линий, варьирующих по продолжительности жизни и по доминирующим причинам смерти. Кроме того, у них не наблюдается энзоотических заболеваний легких, которые усложняют изучение продолжительности жизни у крыс. Различные исследователи изучали специфическое влияние недостаточности калорий и ограничения в потреблении отдельных пищевых продуктов. Танненбаум [1172] обнаружил, что у самок линии ДВА, рост которых был задержан, когда они достигли веса 19-20 г, совершенно отсутствовали спонтанные опухоли, служившие основной причиной смерти у контрольных мышей весом 30 а. У мышей линии YBR/Wi "желтые" богатая жиром диета уменьшает частоту заболеваний амилоидозом, но значительно укорачивает жизнь, а богатая углеводами диета, хотя и вызывает ожирение, не влияет на продолжительность жизни; у мышей линии C57, получавших богатую жиром диету, развивался артрит [1084]. Кинг, Ли и Висчер [599] установили у мышей линии С3Н, которые получали пищу с добавлением обычных микроэлементов, высокую частоту случаев внезапной сердечной недостаточности в возрасте более 1 года, а также рассасывание эмбрионов; эти явления можно было предотвратить, добавляя в пищу токоферол или удаляя из нее микроэлементы. "Тучные" (ob, ob) мыши, увеличение веса которых было задержано путем содержания их на низкокалорийной диете, жили особенно долго - они доживали до 1027 дней [673]; продолжительность жизни у них увеличивалась за счет ограничений в пище больше, чем у "иетучных" гемизиготных особей (ob, + ).
В нескольких случаях лучшие результаты были получены при добавлении витаминов (витамина А, пантотеновой кислоты), но это, вероятно, связано с устранением витаминной недостаточности.
Механизм задержки роста у млекопитающих при недокорме отчасти известен нам по другим работам. Голодание снижает гонадотропную активность гипофиза. Это наглядно показали с помощью трансплантаций Масон и Вольф [771] на самках и Мур и Самуэль [822] на самцах крыс. У крыс уменьшение содержания белков в пище более чем до 7% вызывает анаэструс вследствие снижения гонадотропной активности [450]. При "псевдогипофизэктомии", вызванной недокормом, гипофизарный гормон роста даже при прежнем уровне потребления пищи вызовет возобновление роста скелета и понизит скорость падения веса [838]. Обзор данных по этому вопросу проведен Самуэлем [1034]. Связь между такой "псевдогипофизэктомией" и результатами Мак-Кея до сих пор не ясна, но влияние недокорма на гипофиз, вероятно, играет важную роль в изменении видового возраста.
Карр, Кинг и Висчер [179] создавали у мышей линии СН3 анэструс, уменьшая калорийность пищи вдвое: в возрасте 14 месяцев удавалось легко вызвать единственный цикл введением декстрозы, причем необходимая доза варьировала от 0,15 до 1,0 г. Когда мышам в возрасте 21 месяца стали давать пищу без ограничений, а затем спарили их, все самки забеременели, а 10 особей из взятых в опыт 17 дожили до 23 месяцев и сохранили в этом возрасте половую активность.
У нас нет сравнимых данных для человека. Голодание может вызвать значительную задержку полового созревания (подобно тому или иному заболеванию, ведущему к "косвенному" голоданию, например бильгарциоз); чаще голодание сказывается на всем организме, вызывая уменьшение продолжительности жизни. В экспериментах Мак-Кея ограничение пищевого рациона сводилось к уменьшению его калорийности. Фактически недоедающее большинство не получает от своего образа жизни никакого преимущества в смысле продолжительности жизни. Вполне возможно, однако, как предполагали еще в 1832 г. Эдмондс [319] и впоследствии Синклер [1094] и Мак-Кенс и Уиддоусон [736], что на продолжительности жизни взрослых индивидуумов отрицательно сказывается исключительно быстрый рост в детстве. Наступление половой зрелости у человека можно ускорить путем усиленного питания [151]; получены данные, что хотя максимальный средний рост англичанина за последние 100 лет не увеличился, он достигается по крайней мере на 5 лет раньше [828]; быстрее происходит и уменьшение роста с возрастом. Основные возражения против такого довода состоят в том, что наблюдаемое ускорение полового созревания, вероятно, представляет собой возврат к норме, а не новое приобретение, так как в древние времена менструальный период начинался в том же возрасте, что и сейчас; возникшее "отставание" в начальном периоде развития человека - характерная особенность более "затяжного" детства приматов - вероятно, затруднит любую прямую аналогию с ростом и созреванием грызунов [242, 1173]. Конституциональное преждевременное созревание, по-видимому, не уменьшает продолжительности жизни (ср. фиг. 67).
В 1948 г. Эванс, Симпсон и Ли [330] подтвердили с помощью очищенного гормона роста данные Висчера [1268], что крыс можно держать на протяжении всей жизни в состоянии непрерывного роста, вводя им гипофизарный гормон роста. Висчер отметил некоторое улучшение в состоянии старых самцов крыс под влиянием этого гормона. Цель экспериментов Эванса, Симпсона и Ли заключалась не в изучении влияния роста на продолжительность жизни; эти авторы нашли, что непрерывный рост, вызванный введением крысам гормона, ведет к повышению частоты опухолей и случаев смерти от них. В экспериментах Эванса, Симпсона и Ли 12 животных были забиты на 647-й день для гистологического изучения. При применении малых доз очищенного гормона развивалась устойчивость к действию препарата в отношении ускорения роста и накопления в организме азота [1266]. У собак и кошек непрерывное введение гормона роста после прекращения роста вызывает не рост, а диабет; у других видов (например, у человека)окостенение эпифизарных хрящей предотвращает продолжение роста тела после полового созревания. Реакция крыс на задержку роста и на введение гормона роста, очевидно, является атипичной и, конечно, отличается от реакции, которую можно ожидать у человека; тем не менее имеется возможность сравнивать у млекопитающих влияние на темп старения следующих факторов: 1) задержки роста на все время до достижения видовых размеров, 2) ускорения роста до и после достижения видовых размеров и 3) роста, начинающегося уже в престарелом возрасте и ведущего к превышению видовых размеров. Мун и др. [821] нашли, что введение мышам больших доз гормона роста (2 мг/день) вызывало опухоли только у одной из подопытных линий. Более того, при введении одного только гормона роста не удается вызвать опухоли у гипофиз- эктомированных животных [40, 41]. Невосприимчивость к гетерологичному (бычьему) гормону, использовавшемуся в таких экспериментах, вероятно, можно преодолеть. Идея, лежащая в основе подобных исследований, уже содержалась в работе Робертсона [982, 989], когда эндокринология еще не была настолько развита, чтобы эту идею можно было осуществить. Результаты, полученные при торможении роста "тетелином", почти наверняка не были специфическими. Исследование влияния гормона роста, проведенное на млекопитающих, у которых окостенение эпифизарных хрящей не завершено, можно считать одним из критических экспериментов для выявления того, в какой мере рост и развитие свойственны целостной системе, подверженной старению в определенный конечный момент, и насколько рост, вызванный одним из многих факторов, стимулирующих анаболизм, способен устранить или предотвратить старческие изменения. Эверит [333] пытался выяснить это и нашел, что гормон роста не может ни удлинить жизнь, ни устранить признаки старения у старых крыс.
Попытки ускорить старение у млекопитающих неожиданно оказались безуспешными. Хотя преждевременную гибель лабораторных животных можно вызвать при помощи лекарственных препаратов и ограничений в питании, такие воздействия, за исключением облучения и радиомиметических веществ, не оказывают влияния на процесс старения. Экспериментальные попытки ускорить старение у крыс введением динитрофенола [1167, 1168] или тиреоидина [990] или задержать его тиоурацилом [491] неизменно оказывались неудачными, так как эти препараты не вызывали изменения видового возраста. Петрова [333] показала, что экспериментальный невроз, хотя и не влияет на видовой возраст, приводит к уменьшению продолжительности жизни. Следовательно, у человека наиболее эффективно снизить скорость старческих изменений, ceteris paribus, можно путем улучшения условий жизни в психологическом, социальном и профессиональном отношении.
|
ПОИСК:
|
При использовании материалов проекта обязательна установка активной ссылки:
http://gelib.ru/ 'Геронтология и гериатрия'