3. Гормональная регуляция роста у позвоночных
Для млекопитающих, у которых экспериментально очень трудно расчленить рост и дифференцировку, получено очень много данных относительно старения, в том числе и для очень долгоживущих видов. Для амфибий, у которых имеется четко выраженный метаморфоз и у которых относительно легко можно разделить рост и дифференцировку, до сих пор нет прямых данных относительно старения. Трудно установить, оканчивается ли жизнь интактных амфибий, неотения аксолотля или гигантизм лишенных щитовидной железы головастиков старением по той простой причине, что аксолотли могут жить 50, а нормальные лягушки - 12, 15 или 20 лет. Это осложнение постоянно встречается в исследованиях, посвященных старению. Огромную литературу по эндокринологии и морфогенезу низших позвоночных нельзя использовать при обсуждении данной проблемы ввиду отсутствия статистических данных.
Как гомойотермные, так и пойкилотермные животные независимо от того, есть у них метаморфоз или нет, имеют более раннюю фазу активного роста и более позднюю фазу активного размножения, каждая из этих фаз характеризуется особым типом гормональной регуляции, а для второй фазы характерно относительное ослабление способности к регенерации за счет усиления способности к размножению. Эти фазы различаются по действию механизма, регулирующего скорость процессов, который функционирует в ювенильной фазе. У млекопитающих эти фазы, по-видимому, управляются последовательно гипофизарными механизмами регуляции роста и созревания половых желез. У низших позвоночных процесс дифференцировки и переход к функциям взрослого организма зависит от гипофизарно-тиреоидного равновесия. Гипофизарный гормон роста, выделенный из организма млекопитающих, способен стимулировать рост.
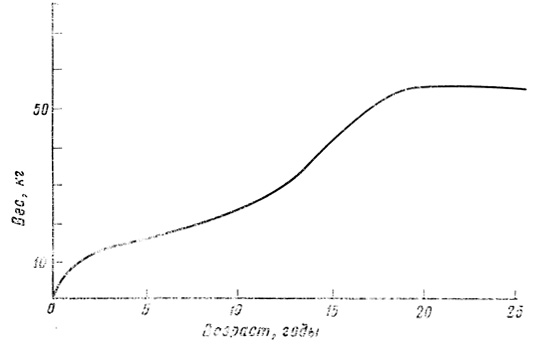
Фиг. 67. Увеличение веса мальчиков с возрастом. По данным Кетле
Для геронтологии особый интерес представляет связь между морфогенезом под влиянием половых гормонов и потерей способности к регенерации. Гробштейн [441] установил, что у рыб из семейства Poecilidae в период дифференцировки гоноподиума под влиянием андрогенов падает способность к регенерации. Он проводит аналогию между этим процессом и потерей способности к регенерации в развивающейся конечности бесхвостых амфибий. Подобные изменения не обязательно связаны с необратимой потерей способности клеток к росту (такая зависимость не проявляется, например, в случае с конечностями амфибий [124, 944]); однако физиологическая способность к регенерации может быть такой же полной, когда речь идет о неповрежденном животном, как и потеря способности к линьке у клопа Rhodnius при утрате эвокатора. Возможно, здесь действует, как считал Майнот, механизм, индуцирующий старческие изменения.
![Фиг. 68. Константа роста у человека (мальчики) по данным Кетле [1048]. К1 и К4 - константы роста для каждого периода](pic/000081.jpg)
Фиг. 68. Константа роста у человека (мальчики) по данным Кетле [1048]. К1 и К4 - константы роста для каждого периода
Имеется некоторое количество данных, касающихся влияния гормонов на анаболизм белка и регуляцию роста у млекопитающих, и особенно у человека. Анализ этих влияний не подтверждает предположения о наличии простой связи между старением и остановкой роста. Еще менее вероятна концепция о единственном, "ведущем" гормональном ингибиторе, который можно изъять из общей программы прогрессирующих изменений в процессе развития. Этой программе, заложенной в человеке, свойственны все сложности динамической системы, в которой гомеостаз сосуществует с изменениями. Большая часть имеющихся сведений носит предварительный характер, и до сих пор не проведено исследований, которые охватывали бы и период старения. Совершенно ясно, однако, что у человека и, вероятно, у некоторых, но не у всех млекопитающих "анаболический" стимул, определяющий синтез белка, не остается одним и тем же на протяжении всей жизни. У взрослого организма он тесно связан с половым циклом. Какова разница в гормональной регуляции роста между организмами с ограниченным (например, человек) и неограниченным (например, крыса) ростом, до сих пор еще не уточнено; крайне мало известно также о гормональной регуляции роста у низших позвоночных. Однако имеющихся данных вполне достаточно для того, чтобы считать неприемлемым любое статическое представление о прекращении роста под влиянием недостаточности ка- кого-то одного гормона. Правильнее, быть может, рассматривать период до достижения половой зрелости и период после ее достижения как отдельные стадии, разделенные чем-то, равнозначным биохимическому метаморфозу.
Рост человека, подобно росту Daphnia (стр. 151), характеризуется двумя перекрывающимися циклами, из которых один относится к периоду до достижения половой зрелости, а другой - к периоду после ее достижения (фиг. 67, 68). Наиболее активная фаза первого цикла приходится на первые 6 месяцев жизни. Этот цикл, согласно Кинселлу [601], почти весь регулируется гипофизарным гормоном роста. Второй цикл, по-видимому, регулируется непосредственно анаболическими стероидными гормонами, выделяемыми половыми железами и корой надпочечников. На протяжении обоих циклов требуется какая-то минимальная секреция гормона щитовидной железы для поддержания роста и развития.
В период половой зрелости в ответ на действие гонадотропного гормона гипофиза половые железы выделяют стероидные гормоны, которые непосредственно стимулируют рост костей и мягких тканей. Процесс роста костей у человека, однако, ограничен, так как одни и те же гормоны обусловливают созревание костной системы и окостенение эпифизарных хрящей. Имеются основания полагать, что эти гормоны подавляют также образование гипофизарного гормона роста - вероятно, через систему отрицательной обратной связи, в которой управляющим сигналом служит интенсивность синтеза белка. Чрезвычайно интересен тот факт, что у мужчин после кастрации иногда наблюдаются симптомы акромегалии [744] или явного гигантизма [575], правда, наряду с полиурией и несахарным диабетом [469]. У девочек после достижения половой зрелости рост, по-видимому, обусловлен в основном деятельностью надпочечников, так как стимулирующее рост влияние эстрогенов и прогестерона менее выражено (за исключением их роли в удержании Са и РО4), по сравнению с соответствующим влиянием андрогенов [601]. Принято считать, что тироксин усиливает у млекопитающих действие гипофизарного гормона роста [331, 1054] в период достижения половой зрелости, а также ускоряет дифференцировку. Это, по всей вероятности, не относится к бесхвостым амфибиям, у которых тиоурацил вызывает ложный гигантизм и для которых, по-видимому, характерно тирео-гипофизарное равновесие [1124]: один гормон вызывает дифференцировку, а другой - рост и пребывание в "ювенильном" состоянии.
Эта картина, требующая детализации, довольно хорошо согласуется с известными данными о роли недостаточности различных гормонов в патогенезе карликовости и гигантизма у человека. Наступление половой зрелости совпадает с завершением созревания костной системы. Введение гормона роста позволяет отсрочить окостенение эпифизарных хрящей и переход к последующему периоду роста [370]; аналогичное явление имеет место и при спонтанном гигантизме. Вместе с тем полное устранение влияния половых желез путем кастрации до достижения половой зрелости не оказывает выраженного влияния на продолжительность жизни; отмечено, что в случаях конституционального преждевременного созревания, когда оно завершается полностью к 6 годам, продолжительность предстоящей жизни, по-видимому, не отличается от нормальной.
Некоторые исследователи считали, что старение млекопитающих сводится к снижению образования гормона роста и обусловлено действием гонадотропного гормона гипофиза на соматические ткани. Если исходить из того, что старение является в сущности результатом дифференцировки, то все сказанное, несомненно, справедливо. Однако в экспериментальном аспекте вопрос заключается скорее в следующем: в какой степени введение одного или нескольких "анаболических" гормонов может изменить у взрослого животного способность поддерживать гомеостаз? Возможно, что гормон роста сам является "ювенильным гормоном" млекопитающего в период до достижения взрослого состояния. Основная его роль заключается в стимуляции белкового синтеза и роста тела. (Отметим, что ювенильный гормон насекомых обнаружен также в органах млекопитающих [1277]; какова его функция там, если он вообще выполняет какую-либо функцию, совершенно неизвестно.) Изменение интенсивности синтеза белков и способности удерживать азот, а также замедление роста представляют собой два явления, наиболее тесно связанные со старением [781]. Введение гормона роста "удивительным образом создает характерные для юношеского возраста соотношения в содержании азота, жиров и воды в организме даже у старых животных" [40, 41], но не увеличивает продолжительность их жизни [333]. Несомненно, изменяется специфичность тканевой реакции на гормон роста; у некоторых млекопитающих это изменение совпадает по времени с достижением зрелости и с вводом в действие нового механизма, поддерживающего анаболизм на постоянном уровне. Экспериментальные исследования Юнга в Англии и Ли в Америке позволяют предполагать, что до какого-то критического момента введение гормона роста лишь стимулирует белковый синтез, а после этого момента он вызывает диабет. Это справедливо для человека, кошек [253] и собак [174], но не для крыс и, по-видимому, не для мышей [821], для которых характерен непрерывный рост. Частота случаев диабета в сочетании со спонтанной акромегалией показывает, что изменение специфичности тканевой реакции связано с влиянием как эндогенного, так и экзогенного гормона. Данные о существовании особого диабетогенного начала не очень убедительны [956, 1297]. Тодоров [1195] с помощью химических методов обнаружил возрастное изменение тканевой реакции на соматотропный гормон даже у крыс; латентный период реакции ДНК и РНК из печени крыс на этот гормон определенно увеличивается с возрастом. Полное удаление передней доли гипофиза у взрослых крыс вызывает прекращение роста; однако больше никаких других данных, которые имели бы отношение к проблеме старения, при этом получить не удается.
В экспериментальных исследованиях высокоочищенный гормон роста при многократном введении крысам уменьшал задержку азота и скорость роста [1266]. Эти эксперименты, однако, неизменно проводились с гетерологичными гормонами (обычно от крупного рогатого скота); поэтому, как в случае антигонадотропного влияния, увеличению реактивности тканей нельзя придавать физиологического значения.
Из других гормонов, оказывающих влияние на процессы роста и дифференцировки, тиреотропный гормон гипофиза у большинства изученных млекопитающих (крысы [1201], кролики [89], мыши [6], крупный рогатый скот [968]) проявляет максимальную активность, по-видимому, в момент наступления половой зрелости; затем происходит снижение активности, которое, однако, никогда не было прослежено до старческого возраста. Снижение интенсивности общего обмена веществ с возрастом, которое часто связывали с уменьшением способности к росту, взятой в качестве показателя "физиологического старения", по-видимому, выражается в ослаблении активности щитовидной железы и, возможно, реактивности клеток, так как после тиреоидэктомии у крыс не наблюдается понижения частоты сердечных сокращений и потребления О2, а у старых нормальных крыс отмечается более слабая реакция на введение тироксина [423]. Снижение интенсивности теплообразования у пожилых людей, возможно, в такой же степени обусловлено инволюцией щитовидной железы, как и атрофией мышц; способность щитовидной железы реагировать на тиреотропин, по-видимому, остается ненарушенной.
Явления, наблюдавшиеся Мак-Кеем при ограничении рациона, можно рассматривать как последствия "алиментарной" гипофизэктомии. Здесь сказываются нарушения в образовании как гормона роста, так и гонадотропного гормона. В результате происходит снижение общей эффективности единой интегрирующей системы роста и развития. Разделение этих систем у млекопитающих представляет собой весьма интересную, но практически трудно разрешимую проблему. Задержка развития путем ограничения рациона значительно отодвигает во времени наступление течки, но все же не может полностью предотвратить ее [35]. Мак-Кей, Сперлинг и Барнес [743] нашли, что задержанные в своем развитии крысы в конце концов утрачивали способность возобновлять рост, если задержка была продолжительной.
Школа Ли и Эванса [40, 41, 238] приступила к разработке проблемы избирательного вмешательства в дифференцировку у млекопитающих. Гипофизэктомия 6-дневных крысят не влияет на прорезывание зубов и прозревание, но подавляет все фазы полового развития. Гипофизэктомированные животные в конце концов умирают от паралича, вызываемого сжатием головного мозга, так как у них мозг растет быстрее, чем черепная коробка. Крысы, выжившие после операции, при введении им гормона роста пребывали в хорошем состоянии. У этих крыс скорость роста была лишь незначительно ниже, чем у контрольных крыс, не подвергавшихся гипофизэктомии. Развитие скелета протекало у них нормально, но характерная для взрослых дифференцировка органов и половое созревание отсутствовали. Наблюдения за тремя такими "метатетелическими" крысами вели в течение 200-300 дней в надежде, что у них разовьется гигантизм. Было бы крайне интересно узнать, какова была бы продолжительность их жизни и какие старческие изменения развились бы у них в конце концов.
Пока еще не удается осуществить избирательное подавление образования гонадотропного гормона, хотя исследования с экстрактами воробейника (Lithospermum) позволяют надеяться на "расчленение" гипофизарного эффекта при помощи химических антагонистов [1270]. Влияние искусственно вызванного преждевременного созревания на продолжительность жизни у млекопитающих (за исключением человека, у которого преждевременное созревание иногда происходит спонтанно), по-видимому, не изучалось. Мышей, у которых созревание наступает очень рано, нельзя считать идеальным объектом для таких исследований, а опыты с более долгоживущими млекопитающими сопряжены с хорошо известными практическими трудностями.
|
ПОИСК:
|
При использовании материалов проекта обязательна установка активной ссылки:
http://gelib.ru/ 'Геронтология и гериатрия'