II. Заключение
На основании приведенных данных трудно дать краткое описание всех влияний излучения на продолжительность жизни. Можно сказать, что излучение, по-видимому, снижает жизненность в том же смысле, в каком гетерозис ее повышает. Сходство между влиянием излучения и инбридинга, подобно сходству между влиянием излучения и старческими изменениями, может быть случайным, но тем не менее заставляет задуматься. У гибридов устойчивость к излучению выше, чем у инбредных организмов [1017]. Излучение, так же как инбридинг и старение, снижает способность организма поддерживать гомеостаз при случайных воздействиях среды. У инбредных животных эта неспособность выражается в сильном размахе изменчивости по таким признакам, как устойчивость к химическим воздействиям [751]; было бы интересно установить при помощи соответствующих тестов, имеется ли подобная изменчивость у облученных животных. Поскольку инбридинг оказывает влияние на протяжении всего развития, инбредные животные, вероятно, в любом случае проявляют большую изменчивость, чем облученные. Но если опыт ставился так, что мышей содержали в условиях постоянного облучения с момента зачатия [846], то они оказывались очень уязвимыми.
Если не считать индукцию опухолей, то излучение не очень сильно влияет на распределение причин смерти - облученные животные, по-видимому, от любой причины умирают несколько раньше. Излучение не ускоряет "развития"; подобно инбридингу, оно чаще замедляет его.
Дальнейшие исследования, возможно, покажут, действительно ли процессы, лежащие в основе этого сходства, идентичны друг другу. Имеется несколько гипотез старения, исходящих из этого сходства и из возможного уменьшения продолжительности жизни за счет облучения [1136]. Старение часто рассматривали как сумму непостоянных изменений в результате повреждений, наносимых внешними факторами, а также побочного действия обмена веществ [385, 582, 1025]. Происходящее с возрастом снижение способности устранять повреждения надо тогда считать следствием дифференцировки и взаимного усиления совпадающих по времени изменений [1025]. Постоянство формы кривой выживания скорее носит статистический характер, но не связано с процессом развития.
Этот динамический вариант старой теории изнашивания (Abnutzungstheorie) будет более приемлемым, если уточнить его, предположив, что изменения затрагивают одну или несколько групп клеток, не способных обновляться путем деления. Ионизирующее излучение может усиливать реактивность структур, которые в норме устойчивы, и таким образом ускорять этот тип изнашивания (подобно тому как песок повышает скорость износа подшипников), вызывая те же изменения на молекулярном уровне [483]. Возникает возможность непосредственно определить как продолжительность жизни молекул в клетках, так и влияние на нее излучения у животных с постоянным числом клеток. Столько же "почтенные" теории изнашивания клеточных и неклеточных микроструктур [763, 764, 1023] (если не как причины старения, то хотя бы как сопутствующего явления) нашли некоторое подтверждение в последних работах по коллагену [62, 1225]. Однако в этом случае излучение не приводит к ускорению процесса [1095]. Быть может, сходство между процессами старения и лучевого повреждения касается процессов восстановления, а не природы повреждений [1135].
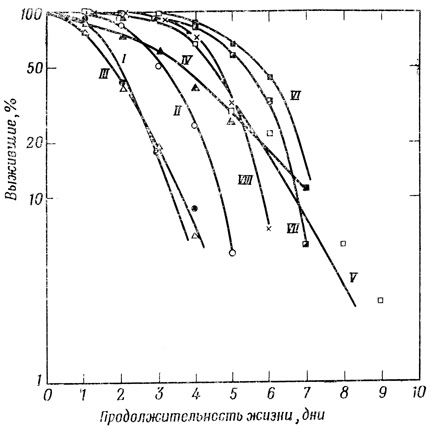
Фиг. 72. Кривые выживания гидрантов Campanularia plexuosa после воздействия рентгеновскими лучами. I - контроль, 23 особи, средняя продолжительность жизни 2,7±0,3 дня; II - 500 р, 81 особь средняя продолжительность жизни 3,6±0,07 дня; III - 2000 р, 49 особей, средняя продолжительность жизни 2,41±07 дня; IV - 5000 р, 15 особей, средняя продолжительность жизни 3,87±0,3 дня; V - 25 000 р, 37 особей, средняя продолжительность жизни 4,88±0,25 дня; VI - 50 000 р, 9 особей, средняя продолжительность жизни 6,11 ±0,4 дня; VII - 100 000 р, 51 особь, средняя продолжительность жизни 5,78±0,17 дня; VIII - 200 000 р, 15 особей, средняя продолжительность жизни 5,07±0,22 дня
Весьма интересные результаты получил Стрелер [1328] на Campanularia (кишечнополостные). Продолжительность жизни отдельных гидрантов в норме составляет 2-3 дня. После облучения же (при дозе до 50 000 р) она значительно увеличивалась и даже при дозе 200 000 р возрастала вдвое (фиг. 72). Никакого объяснения этим уникальным данным не предложено. Возможно, под влиянием облучения выводится из строя существующий или запрограммированный механизм умирания.
Вполне естественно, что основное внимание радиобиологов было устремлено на изучение влияния излучения на хроматин. На сходство кривых смертности и кривых изменения энтропии указывал еще в 1825 г. Гомпертц. Перке [929] отметил, что уменьшение жизненности, измеряемое как "неспособность противостоять разрушению", имеет ту же природу, что и "рассеяние энергии", т. е. увеличение энтропии, которая служит мерой неупорядоченности системы. В настоящее время представляется вполне вероятным, что информация, содержащаяся в клетках, и есть та "биологическая энергия", существование которой предполагали в прошлом столетии и о которой думали, что она растрачивается с возрастом. Инбридинг может уменьшать объем информации, ограничивая разнообразие имеющегося генетического материала [184, 422, 788]. Излучение может осуществлять это, оказывая специфическое действие (точковая мутация, разрывы хромосом) или просто повышая уровень шума в системе; старение может отражать сходное повышение уровня шума вследствие метаболического "изнашивания" соответствующих молекул и случайных поражений. Математический анализ таких представлений [1025] опередил биологический эксперимент. Их не удается еще связать, например, с нормальными межвидовыми различиями в продолжительности жизни. Некоторые, если не большинство радиационных поражений клеток, по крайней мере у ресничных инфузорий, носят экзогенный характер. Одной только точковой мутации, вероятно, недостаточно для статистического объяснения наблюдаемых темпов старения животных [334]. Рассел [1018] показал, однако, что уменьшение продолжительности жизни у самцов мышей, подвергшихся облучению при взрыве атомной бомбы, передается их потомству. Особенно интересны данные Гармана [483], что противолучевые препараты увеличивают продолжительность жизни у необлученных мышей.
Гипотермия, подобно недокорму, также создает некоторую защиту от излучения [529]. Правда, рост облученных крыс, получавших ограниченный рацион, при переводе их на нормальное питание не возобновляется [182]. Получение таких результатов, однако, зависит главным образом от строгого соблюдения методики эксперимента; лишь теперь стало ясно, как трудно их соблюдать. Установлено, однако, что полиплоиды необычайно устойчивы к излучению [1115], а гаплоиды (наездник Habrobracon [206, 207]) более чувствительны, чем диплоиды: под влиянием облучения продолжительность жизни у них уменьшается в большей степени. Кертис и Гебгард [266] нашли, что токсичные вещества неспецифического действия и азотные аналоги иприта не ускоряют процесс старения, но позднее появились сообщения, что химические мутагены приводят к укорочению жизни [12, 249, 1211]. Эти последние данные устраняют одно серьезное возражение против мутационной теории старения.
От радиационной биологии следует, вероятно, ждать серьезного вклада в понимание процессов естественного старения. Наиболее перспективным в этом отношении исследованием, не требующим никаких допущений об идентичности радиационного поражения и возрастных изменений, будет, по нашему мнению, изучение природы жизненности. Как только будут поняты клеточные и физиологические последствия изменений жизненности под влиянием облучения, станут ясны и другие возможные области применения радиационной биологии. По крайней мере в настоящее время нет других подходов к изучению проблемы старения, которые были бы более перспективными. Геронтология и радиационная биология, по-видимому, примерно в равной мере способствовали пониманию отдаленных последствий облучения. Любая современная программа в области здравоохранения должна содействовать проведению исследований в этих двух направлениях. Достаточно много времени было уделено экспериментам по действию излучения на животных, в которых показано, что излучение ничего хорошего им не приносит. Поскольку мы в этом убедились, основное внимание следовало бы направить на более поучительные сравнения.
Имело бы смысл сравнить действие низких доз излучения на животных со строго постоянным и непостоянным числом клеток. Имело бы также смысл исследовать влияние частичного облучения тела и отдельных органов с применением современных методов ортотропных трансплантаций (или без них). Имеются также прекрасные возможности для относительно простого сравнения реакции гибридных или инбредных линий и взятых от них клеток. Для понимания старения тканей важнее всего разработать методы анализа эпигенетического изменения клеток на протяжении всей жизни, их обновления и влияний дифференцировки, а также методы исследования ядерных и цитоплазматических структур. Без предшествующих достижений в этой области нельзя достигнуть реального и детального понимания действия облучения на жизненность млекопитающих.
"Наиболее рациональное и единственное объяснение функционального поражения стареющей ткани заключается в том, что оно отражает абсолютную утрату клеток в популяции с замещением их внеклеточной жидкостью или соединительной тканью; в этом суть гипотезы изнашивания клеток" [477]. Очень трудно, однако, по вышеприведенным соображениям связать этот процесс с простой "генетической смертью" (в результате мутации) неизменного числа постмитотических клеток. Еще труднее связать его с мутационной теорией уменьшения продолжительности жизни под влиянием облучения. Имеющиеся данные позволяют предполагать, что при дозе ниже 200 р у млекопитающих повреждаются именно хромосомы (при средней дозе, вызывающей разрыв хромосом, 19 рад). Чувствительность клеток млекопитающих в культуре была в основном подтверждена и in vivo. Средняя летальная доза для клеток человека в интерфазе оказалась равной примерно 86 рад. Все имеющиеся данные свидетельствуют о том, что чувствительность неделящихся клеток весьма вариабельна- [516, 950, 1193]: у животных, у которых нет клеточного деления, таких, как наездник (Habrobracon), излучение действительно вызывает уменьшение продолжительности жизни, причем у гаплоидных особей в большей степени, чем у диплоидных, но только при дозах, превышающих в 10 или 100 раз дозы, которые сокращают жизнь грызунов [206, 207]. Кроме того, имеются указания на то, что реактивность клонально делящихся клеток изменяется с возрастом и под влиянием малых доз излучения. Прат установил, что у облученных мышей наблюдается такой же лаг-период в процессе восстановления печени после гепатэктомии, как и у старых мышей [950]. Даже если вместо толковой мутации взять повреждение хромосом как более обычный механизм старения и радиационного повреждения (сделав допущение, что их механизмы одинаковы, что, возможно, и неверно), создается впечатление, что выживание поврежденных, но клонально жизнеспособных клеток играет более важную роль в уменьшении жизненности и нарушении гомеостаза, чем утрата клеток [950]. Утрата неделящихся клеток может быть вторичной; если же она первична, то мы не можем сказать, какой из этих двух механизмов определяет продолжительность жизни.
|
ПОИСК:
|
При использовании материалов проекта обязательна установка активной ссылки:
http://gelib.ru/ 'Геронтология и гериатрия'