3.4. Эпидемиологический подход к изучению биологии продолжительности жизни человека
Среди методов изучения биологических основ продолжительности жизни человека эпидемиологическим исследованиям принадлежит особая роль, поскольку возможности постановки экспериментов на человеке ограничены для ученых, но безграничны для природы. Эпидемиология, зародившись как раздел медицинской географии, изучающий очаги инфекционных болезней и закономерности развития эпидемий, впоследствии необычайно расширила круг рассматриваемых проблем. Началось ускоренное и успешное развитие эпидемиологии неинфекционных заболеваний, и в том числе эпидемиологии рака [Долл, Пито, 1984]. Традиционные методы эпидемиологических исследований, основанные на сопоставлении различных географических районов, дополнились анализом особенностей разных групп населения, живущих на одной и той же территории. В результате современная эпидемиология переросла рамки медицинской географии и чисто медицинской проблематики, превратившись в мощный инструмент разработки фундаментальных медико-биологических проблем. Одна из них - выяснение биологических основ продолжительности жизни человека.
До последнего времени эпидемиологические исследования проблемы продолжительности жизни ограничивались в основном анализом феномена долгожительства [Зубов, Козлов, 1982]. Такой подход имеет, однако, два существенных недостатка. Во-первых, возраст долгожителей постоянно преувеличивается [Abrams, 1985], поэтому неясно, что же исследуется: эпидемиология долгожительства или эпидемиология легенд и анекдотов. Во-вторых, география очагов долгожительства может быть обусловлена не современными условиями, а теми неизвестными сегодня обстоятельствами, которые существовали много лет назад. Поэтому значительно более перспективным представляется анализ надежных демографических данных по смертности взрослых людей (20-80 лет) с целью выявления биологических характеристик продолжительности жизни и последующего изучения региональных различий.
Вместе с тем поиск фундаментальных биологических различий в смертности человеческих популяций осложняется мощным влиянием социальных факторов. Поэтому само по себе сопоставление уровней смертности людей, живущих в разных социально-экономических условиях, мало что может дать для выявления биологических основ продолжительности жизни. Теоретически эту проблему можно было бы решить, учитывая влияние каждого из мешающих факторов, с тем, чтобы вычитая их влияние, получить биологически обусловленные остаточные различия. На практике, однако, такой метод исключения представляет собой почти безнадежную задачу, поскольку трудно даже перечислить все возможные мешающие факторы, не говоря уже о количественном учете их влияния. По этой причине эпидемиологические исследования биологии продолжительности жизни человека развивались чрезвычайно медленно.
Ситуация может принципиально измениться теперь, когда найдены описанные в предыдущем разделе биологические характеристики продолжительности жизни человека. Как уже отмечалось, анализируя возрастную динамику смертности людей, можно выделить такую компоненту смертности, которая остается исторически стабильной, несмотря на резкое снижение общей смертности и радикальные социально-экономические преобразования, происшедшие в XX в. Историческая стабильность этой компоненты смертности указывает на то, что она обусловлена фундаментальными биологическими (генетическим и экологическими) особенностями популяций человека, которые эволюционируют значительно медленнее, чем социально-экономические условия жизни. Таким образом, открывается уникальная возможность изучать биологически обусловленные различия по продолжительности жизни людей на сложном фоне социально-экономических различий сравниваемых популяций.
Перед тем как переходить к анализу региональных различий по биологическим характеристикам продолжительности жизни, необходимо решить вопрос о том, какие именно группы населения следует сопоставлять. И прежде всего возникает проблема, как анализировать данные по смертности мужчин и женщин - совместно или раздельно? Действительно, между этими двумя большими группами населения обычно существуют значительные различия в уровнях смертности (превышение мужской смертности над женской). С другой стороны, если такие различия имеют в основном социальную природу, как полагают некоторые демографы, то биологические характеристики продолжительности жизни обоих полов должны быть близки друг к другу. Вместе с тем опыт предыдущих исследований (см. историю расчета C-критерия в разделе 3.2) показал, что совместный анализ данных по мужчинам и женщинам нередко является чрезмерным упрощением. Поэтому прежде всего следует выяснить, с какими параметрами распределения продолжительности жизни связаны различия в смертности мужчин и женщин.
Сравнительный анализ распределения продолжительности жизни мужчин и женщин. Для изучения половых различий в смертности людей был использован метод попарных сопоставлений для каждого из трех параметров формулы Гомперца-Мейкема. Всего было сделано 255 попарных сравнений для каждого параметра в географическом и историческом разрезах. Оказалось, что величина фоновой компоненты интенсивности смертности (параметр A) мужчин и женщин была, как правило, одинаковой в пределах ошибки измерений. Если же сопоставлять эти данные в целом, то отсутствует даже тенденция к каким-либо различиям по этому параметру: в половине случаев оценка параметра А для женщин несколько превышала соответствующую оценку для мужчин (53±6% для P≥0,95), а в оставшейся половине случаев (47±6% для P≥0,95) наблюдалась обратная картина. Следовательно, половые различия в продолжительности жизни связаны не с фоновой, а с возрастной компонентой интенсивности смертности. Действительно, оказалось, что возрастная компонента интенсивности смертности мужчин, как правило, достоверно превышает возрастную интенсивность смертности женщин. Более того, эти различия образуют достоверную тенденцию при тотальном сопоставлении данных: в 97,6% случаев (95,2-99,0% для P≥0,95) оценка возрастной компоненты интенсивности смертности 40-летних мужчин превышала соответствующую оценку для женщин. Выявленная закономерность обусловлена различиями как по параметру R, так и по параметру α. Оценка R для мужчин в 94,7% случаев (91,5-97,1% для P≥0,95) превышала соответствующую оценку для женщин. Для параметра а наблюдалась обратная картина: в 92,3% случаев (88,7-95,3% для P≥0,95) оценка а у женщин была выше. Таким образом, возрастная компонента интенсивности смертности и период ее удвоения у женщин меньше, чем у мужчин. Табл. 6 иллюстрирует обнаруженные закономерности на конкретных примерах. В этой таблице приведены медианные значения параметров распределения и возрастной интенсивности смертности для 40-летнего возраста, а также доверительные интервалы для этих медианных значений (приведены в скобках и соответствуют доверительной вероятности 97-99%). Материалы таблицы получены методами непараметрической статистики [Campbell, 1967], поскольку распределение оценок параметров могло отличаться от нормального. Можно заметить, что половые различия продолжительности жизни ярко выражены в Финляндии и во Франции, а в Швеции - минимальны.
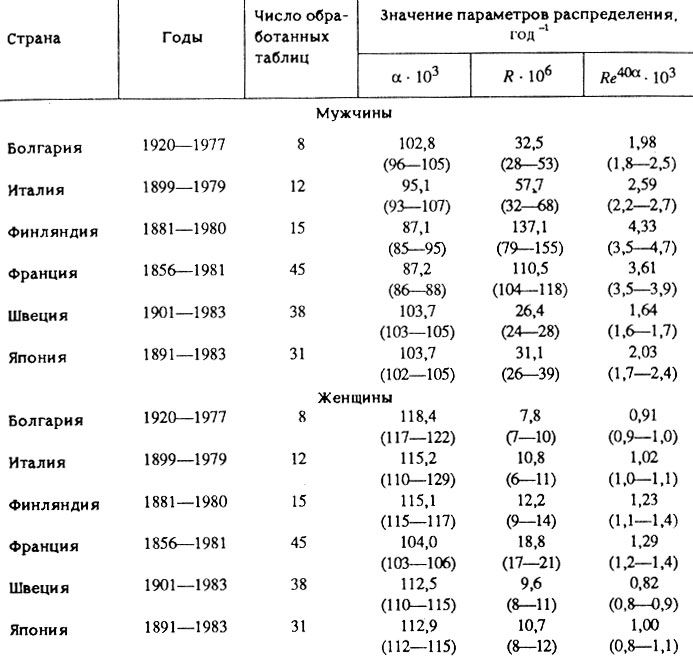
Таблица 6. Сравнительная характеристика распределения продолжительности жизни мужчин и женщин
Таким образом, половые различия в смертности взрослых мужчин и женщин связаны в основном с исторически стабильными (биологическими) характеристиками продолжительности жизни человека. Следовательно, анализ региональных различий по этим характеристикам необходимо проводить раздельно для мужской и женской частей населения.
Эпидемиология биологических характеристик продолжительности жизни человека. Как видно из приведенной выше табл. 6, величина возрастной компоненты смертности, будучи исторически стабильной, сильнейшим образом зависит от пола и проявляет значительную региональную изменчивость.
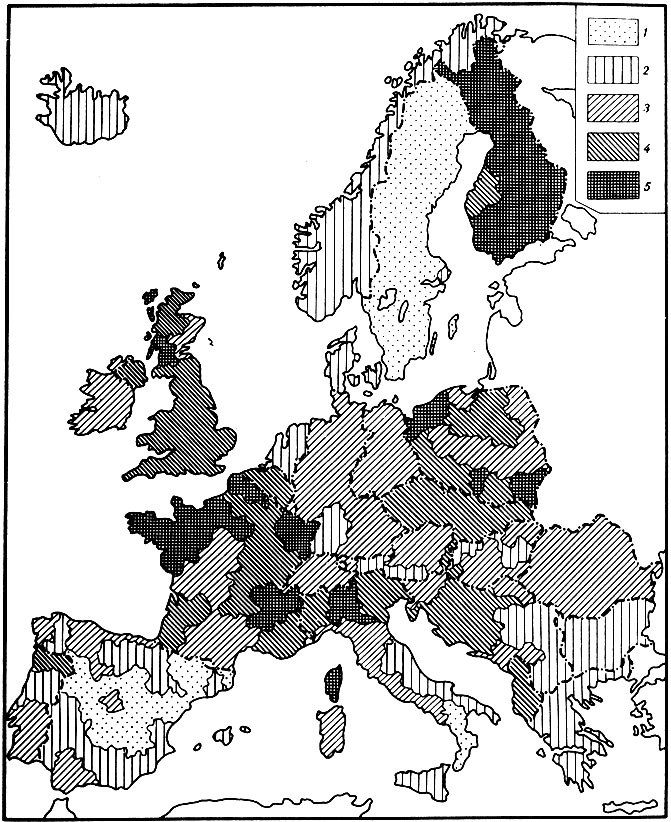
Рис. 13. Картограмма уровней возрастной (биологической) компоненты смертности мужчин для возраста 40 лет на территории зарубежной Европы. 1 - низкий уровень, менее 1,6*10-3 год-1; 2 - пониженный уровень: (1,61- 2,20)*10-3 год-1; 3 - средний уровень: (2,21-2,80)*10-3 год-1; 4 - повышенный уровень:(2,82-3,40)*10-3 год-1; 5 - высокий уровень: свыше 3,41*10-3 год-1
На рис. 13 и 14 приведены картограммы уровней биологической (возрастной, исторически стабильной) компоненты смертности мужского и женского населения зарубежной Европы в возрасте 40 лет. В дальнейшем мы покажем, что выбор того или иного возраста влияет только на абсолютную величину биологической компоненты смертности, не меняя рангового положения районов по величине данного показателя (см. раздел 4.5). Следует отметить, что уровень биологической компоненты смертности женщин оказался настолько ниже соответствующего уровня для мужчин, что при построении картограмм пришлось использовать разную абсолютную шкалу градаций уровней этого показателя.
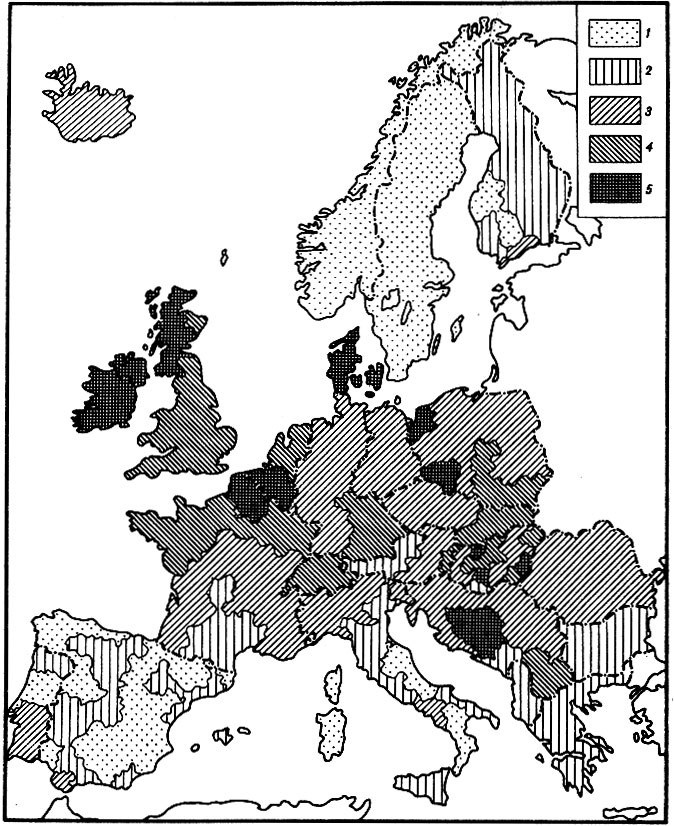
Рис. 14. Картограмма уровней возрастной (биологической) компоненты смертности женщин для возраста 40 лет на территории Зарубежной Европы. 1 - низкий уровень: менее 0,83*10-3 год-1; 2 - пониженный уровень: (0,84-1,03)*10-3 год-1; 3 - средний уровень: (1,04 -1.23)*10-3 год-1; 4 - повышенный уровень: (1,24-1,43)*10-3 год-1; 5 - высокий уровень: свыше 1,44*10-3 год-1
Сопоставляя картограммы, можно заметить, что между ними имеется определенное сходство. С одной стороны, в обоих случаях низкие уровни биологической компоненты смертности наблюдаются в Швеции, Норвегии, Греции, Болгарии, Испании и на юге Италии. С другой стороны, северо-западная часть Европы (Великобритания, север Франции и Бельгия) характеризуются высоким уровнем биологической компоненты смертности как для мужчин, так и для женщин.
Вместе с тем наряду со значительным сходством можно отметить также существование некоторых различий между картограммами. Так, например, мужское население Финляндии характеризуется самым высоким уровнем биологической компоненты смертности в Европе. В то же время женское население Финляндии занимает промежуточное положение по изучаемому признаку.
Сопоставление этого факта с другими эпидемиологическими данными подтверждает особое положение мужского населения Финляндии. Действительно, известно, что один из самых высоких в мире уровней смертности от сердечно-сосудистой патологии наблюдается в Финляндии, причем в первую очередь среди мужчин. Более того, особенно высокая смертность от этого вида патологии зарегистрирована именно в восточных районах Финляндии (Северная Карелия) [Salonen et al., 1982], где, согласно нашим расчетам, наблюдается также и наиболее высокий уровень биологической компоненты смертности. Таким образом, обнаруженная закономерность (высокий уровень биологической компоненты смертности у мужчин Финляндии) получает независимое подтверждение из других эпидемиологических исследований.
Обсуждая возможные механизмы этого явления, следует отметить, что у мужчин Финляндии содержание холестерина в крови одно из самых высоких в мире [Knuiman et al., 1982]. При этом особенно высокое содержание холестерина зарегистрировано у мужчин, живущих в восточной части Финляндии (6,3-6,4 ммоль/л), где наблюдается самый высокий уровень биологической компоненты смертности [Гаврилов, Гаврилова, 1986] и смертности от болезней сердца [Salonen et al., 1982]. У мужчин западной части Финляндии содержание холестерина в крови оказывается несколько ниже (5,9-6,2 ммоль/л), но все равно значительно превышает уровень, характерный для большинства других стран (так, например, у мужчин Италии концентрация холестерина составляет 5,1-5,3 ммоль/л) [Knuiman et al., 1982].
Значение этих фактов очевидно, если учесть, что высокое содержание холестерина в крови считается важным фактором риска возникновения сердечно-сосудистой патологии [Knuiman et al., 1982].
Другая интересная особенность Финляндии состоит в том, что в почвах и водах этой страны содержится необычайно мало микроэлемента селена [Mutanen, Koivistoinen, 1983]. В результате концентрация селена в крови у финнов оказывается значительно ниже уровней, характерных для других стран. Так, например, содержание селена в крови у жителей г. Хельсинки составляет всего 0,081 мкг/мл, что почти в три раза меньше, чем у населения большинства других стран мира. Для сравнения укажем, что средний уровень содержания селена в крови населения США составляет 0,206 мкг/мл, а у жителей Венесуэлы - 0,355 мкг/мл [Levander, 1982]. Интересно отметить, что особенно низкий уровень селена в крови наблюдается опять-таки у населения восточных районов Финляндии. Например, у жителей г. Лаппенранта (Северная Карелия) содержание селена составляет всего 0,056 мкг/мл [Levander, 1982].
Обсуждая биологические последствия недостатка селена в организме, необходимо иметь в виду следующие факты. Известно, что недостаток селена в диете экспериментальных животных приводит к возникновению сердечной патологии и ряда других расстройств [Levander, 1982]. Эпидемиологические исследования подтвердили, что в районах с низким содержанием селена наблюдается повышенная смертность от целого ряда заболеваний, включая сердечно-сосудистые [Shambergcr et al., 1978]. Более того, в ходе этих эпидемиологических исследований выяснилось, что мужское население особенно чувствительно к дефициту селена. Так, например, оказалось, что коэффициент корреляции между смертностью от болезней органов кровообращения и содержанием селена в крови для населения 19 штатов США составляет -0,79 для мужчин и только -0,53 для женщин. Аналогичные коэффициенты корреляции для смертности от гипертонической болезни составляли -0,84 для мужчин и всего -0,54 для женщин. [Shamberger et al., 1979]. Более того, оказалось, что недостаток селена повышает риск возникновения злокачественных новообразваний у мужчин почти в 3 раза, но не влияет существенно на риск возникновения рака у женщин [Kok et al., 1987]. Роль недостатка селена в возникновении болезней органов кровообращения была подтверждена и в ходе длительного проспективного исследования, проведенного в районах Финляндии с низким содержанием селена [Salonen, 1987]. Выяснилось, что у мужчин с содержанием селена в сыворотке крови менее 0,045 мкг/мл риск смерти от сердечно-сосудистых заболеваний повышен в 2,7 раза, а риск смерти от ишемической болезни сердца - в 3,6 раза.
Таким образом, обсуждаемая "селеновая" гипотеза позволяет объяснить целый ряд фактов: высокий уровень биологической смертности мужчин в Финляндии (особенно в районах с низким содержанием селена), преимущественное поражение мужской части населения и характерный тип патологии (болезни органов кровообращения).
Дальнейшее более детальное обсуждение роли селена в жизнедеятельности организмов выходит за рамки этой книги. Отметим лишь, что селен является составной частью фермента глютатионпероксидазы, входя в активный центр этого фермента в виде селено-цистеина [Levander, 1982]. Этот фермент восстанавливает перекись водорода до воды, а органические гидроперекиси - до соответствующих спиртов, препятствуя таким образом перекисному окислению биологических мембран и повреждению других клеточных структур [Harman, 1981]. Недостаток селена приводит к уменьшению активности данного фермента, что способствует развитию целого ряда патологических процессов, включая болезни сердца и ускоренное старение. Повышенная устойчивость женщин к недостатку селена связана, по-видимому, с тем, что женский организм более защищен от повреждения свободными радикалами [Harman, 1981; 1985].
Завершая обсуждение "селеновой" гипотезы, следует отметить, что в 1969 г. вошло в силу Постановление Министерства сельского хозяйства и лесоводства Финляндии о добавлении селена в корм скоту, что должно было привести к повышению содержания данного микроэлемента в мясо-молочных продуктах [Mutanen, Koivistoinen, 1983]. Поэтому если "селеновая" гипотеза верна, то можно было бы ожидать снижения биологической компоненты смертности мужского населения Финляндии. Действительно, в 70-х годах произошло некоторое повышение содержания селена в крови у финнов [Salonen, 1987], которое сопровождалось снижением общей смертности и смертности от коронарной болезни сердца [Werko, 1987]. Следует особо отметить, что наблюдаемое снижение смертности не удается объяснить изменением общеизвестных факторов риска [WerkO, 1987], что заставляет более внимательно отнестись к "селеновой" гипотезе. Обращает на себя внимание также и то, что именно в 70-х годах, как показали наши расчеты [Гаврилова. 1982], у мужчин Финляндии началось уменьшение биологической компоненты смертности. В 1976-1980 гг. уровень этой компоненты для 40-летних мужчин составил 79% от уровня, наблюдавшегося в 1966-1970 гг.
Приведенный пример показывает, что разделение смертности на социальную и биологическую компоненты в значительной мере является условным и исторически преходящим. Фактически биологической компонентой является та часть смертности, с которой человек еще не научился бороться. Можно не сомневаться, что по мере дальнейшего развития науки все большую часть смертей можно будет поставить под социальный контроль и устранить. Тем не менее разделение смертности на социальную и биологическую компоненты оказывается полезным для решения данной задачи.
Другой пример несовпадения картограмм уровней биологической компоненты смертности для мужчин и женщин касается Дании. Можно заметить, что мужское население этой страны характеризуется сравнительно низким уровнем биологической компоненты смертности. В то же время женское население Дании имеет один из самых высоких в Европе уровней этого показателя. Необходимо, правда, отметить, что по абсолютной величине уровни биологи-ческой смертности мужчин и женщин Дании соизмеримы. Иначе говоря, наиболее высокий уровень биологической смертности женщин оказывается все же несколько меньше, чем наиболее низкий уровень биологической смертности мужчин. Тем не менее возникает вопрос, с чем связана столь высокая биологическая смертность женщин Дании по сравнению с женским населением других европейских стран.
В связи с этим интересно отметить, что данное наблюдение также хорошо согласуется с материалами эпидемиологических исследований, проведенных ранее. Оказывается, женское население Дании характеризуется одним из самых высоких в мире уровней онкологической заболеваемости и смертности, включая смертность от рака молочной железы. Причины этого явления до сих пор не установлены, однако известно, что эта страна традиционно отличается одним из самых высоких в мире уровнем потребления жиров. Этот уровень в стране составляет 170 г в день на человека, что почти на 40% больше, чем в Финляндии, и почти в шесть раз больше, чем в Японии [Долл. Пито. 1984]. Многочисленные эпидемиологические исследования [Knox, 1977; Долл, Пито, 1984; Hopkins, Carroll, 1985], а также эксперименты на животных [Hopkins, Carroll, 1985] показали, что избыточное потребление жиров может быть важным фактором развития целого ряда заболеваний и способствовать развитию старческих изменений [Harman, 1981]. Разумеется, данное объяснение является лишь гипотезой, иллюстрирующей возможность применения этих картограмм для выяснения биологических основ продолжительности жизни человека.
Завершая обсуждение картограмм, следует обратить внимание на величину наблюдаемых различий. В целом для женского населения Европы характерна меньшая вариабельность по уровню биологической компоненты смертности. Так. для возраста 40 лет различия между наилучшими и наихудшими районами составляют 200% у женщин и 300% у мужчин. Это очень большие различия, особенно если учесть, что выраженной тенденции к их уменьшению не наблюдается. Поэтому выяснение причин наблюдаемых региональных различий позволило бы снизить смертность трудоспособного населения в два-три раза, что имеет не только научное, но и большое практическое значение. В этом отношении значительный интерес представляет изучение районов, где происходят "скачки" смертности от самого низкого к самому высокому уровню, например пограничные районы Финляндии с Норвегией и Швецией (для мужчин) или Дании с Норвегией и Швецией (для женщин). Разумеется, особый интерес представляют такие скачки смертности в пределах соседних районов одной и той же страны, характеризующихся сходными условиями жизни. Перспективным представляется изучение смертности мигрантов, которое позволило бы различать влияние генетических и средовых факторов на биологическую компоненту смертности.
Несмотря на все разнообразие возможных гипотез, объясняющих наблюдаемые региональные различия, их можно свести к четырем обобщенным гипотезам: гипотезе средовой детерминации, гипотезе генетической детерминации, гипотезе социокультурной детерминации и гипотезе адаптационной детерминации.
Согласно гипотезе средовой детерминации, биологическая компонента смертности определяется исторически стабильными факторами окружающей среды. Примером такой гипотезы является обсуждавшееся ранее предположение о роли дефицита селена в повышении биологической компоненты смертности мужчин Финляндии. В соответствии с гипотезами данного типа следует ожидать, что биологическая компонента смертности мигрантов должна определяться новым местом их проживания.
Согласно гипотезе генетической детерминации, биологическая компонента смертности наследуется так же. как и многие другие расовые признаки. Если эта гипотеза верна, то биологическая компонента смертности мигрантов должна определяться в основном их этнической и расовой принадлежностью.
Согласно гипотезе социокультурной детерминации, биологическая компонента смертности определяется такими коренными чертами образа жизни, как. например, диета и ряд специфических национальных привычек. Здесь следует подчеркнуть, что речь идет о таких социокультурных особенностях, которые настолько устойчивы, что их можно рассматривать как компоненты экологии человеческих популяций. К этой гипотезе относится уже высказанное предположение о роли высокого потребления жиров в Дании как фактора высокой биологической компоненты смертности женщин. В соответствии с гипотезой социокультурной детерминации следует ожидать, что у ассимилировавшихся мигрантов уровень биологической смертности должен определяться новой средой обитания, а у мигрантов, сохранивших национальные черты и привычки,- остаться неизменным.
Наконец, гипотеза адаптационной детерминации предполагает, что биологическая смертность определяется степенью адаптированности человеческой популяции к среде своего обитания. Гипотеза предсказывает, что биологическая смертность мигрантов должна быть, как правило, выше, чем в районах, откуда они прибыли, и по сравнению с коренным населением в районе прибытия. Согласно этой гипотезе, следует также ожидать чрезвычайно низких уровней биологической компоненты смертности у народов, веками живущих на одном и том же месте. В этой связи интересно отметить, что в таких странах, как Швеция и Норвегия, наблюдается низкий уровень биологической компоненты смертности, несмотря на то что климатические условия в этих странах не являются особенно благоприятными. Данную гипотезу можно рассматривать как частный случай представления об адаптивных типах людей, возникших в результате длительной истории приспособления человеческих популяций к различным условиям обитания [Алексеева, 1986].
Разумеется, эти гипотезы не исключают, а скорее дополняют друг друга. Ясно также, что изучение биологической компоненты смертности мигрантов является одним из наиболее перспективных направлений дальнейших исследований эпидемиологии биологических характеристик продолжительности жизни человека.
Достоверность обнаруженных региональных различий. Хотя приведенное ранее обсуждение картограмм носило фрагментарный и отчасти спекулятивный характер, ясно видно, что подобный эпидемиологический подход в принципе позволяет исследовать биологические основы продолжительности жизни человека. Поскольку научное и практическое значение таких исследований трудно переоценить, еще раз вернемся к вопросу о достоверности обнаруженных региональных различий.
Под достоверностью в данном случае мы понимаем не статистическую значимость региональных различий, в которой легко убедиться, обратившись, например, к табл. 6, а правильность конечных результатов, основанных хоть и на проверенных, но не абсолютно строгих допущениях (справедливость формулы Гомперца-Мейкема. историческая стабильность возрастной компоненты интенсивности смертности). Иначе говоря, перед тем как углубляться в изучение природы обнаруженных различий, полезно еще раз независимым способом проверить, действительно ли существуют эти различия, или они являются артефактом использованного метода обработки данных.
Необходимо также отметить, что сам факт существования биологических различий в смертности человеческих популяций является неочевидным. Например, некоторые демографы полагают, что все люди исходно равны в отношении своей возможной продолжительности жизни, а существующие региональные различия в смертности связаны исключительно с влиянием социальных факторов. Поскольку полученные результаты противоречат этому весьма распространенному мнению, то следует с особой тщательностью проверить их достоверность.
Для проверки реальности существования обсуждаемых различий посмотрим, можно ли на их основании делать правильные и нетривиальные предсказания. Ниже приведено несколько примеров таких наиболее парадоксальных прогнозов смертности и их проверки.
![Рис. 15. Сопоставление исторической динамики смертности женщин Норвегии и Дании 1 - общая интенсивность смертности женщин Норвегии в возрасте 40 лет: 2 - общая интенсивность смертности женщин Дании в возрасте 40 лет; 3 - возрастная компонента смертности женщин Норвегии в возрасте 40 лет; 4 - возрастная компонента смертности женщин Дании в возрасте 40 лет. Рассчитано и построено на основании статистических данных [см.: Гаврилова и др., 1983]](pic/000113.jpg)
Рис. 15. Сопоставление исторической динамики смертности женщин Норвегии и Дании 1 - общая интенсивность смертности женщин Норвегии в возрасте 40 лет: 2 - общая интенсивность смертности женщин Дании в возрасте 40 лет; 3 - возрастная компонента смертности женщин Норвегии в возрасте 40 лет; 4 - возрастная компонента смертности женщин Дании в возрасте 40 лет. Рассчитано и построено на основании статистических данных [см.: Гаврилова и др., 1983]
Первый пример относится к сопоставлению смертности женщин Дании и Норвегии. В начале XX в. смертность женщин в Норвегии была существенно выше, чем в Дании (рис. 15) без каких-либо признаков того, что эта ситуация должна измениться. Однако из тех же таблиц дожития, относящихся к началу века, было рассчитано, что биологическая (возрастная) компонента смертности в Норвегии не больше, а, наоборот, существенно меньше, чем в Дании (рис. 15). Поэтому следовало ожидать, что по мере улучшения условий жизни и уменьшения социальной (фоновой) компоненты смертности должна произойти историческая инверсия смертности - Норвегия и Дания должны поменяться местами по уровню смертности женщин. Как видно из рисунка, это предсказание полностью подтвердилось.
![Рис. 16. Сопоставление исторической динамики смертности женщин Болгарии и Италии. 1 - общая интенсивность смертности женщин Болгарии в возрасте 40 лет; 2 - общая интенсивность смертности женщин Италии в возрасте 40 лет; 3 - возрастная компонента смертности женщин Болгарии в возрасте 40 лет: 4 - возрастная компонента смертности женщин Италии в возрасте 40 лет. Рассчитано и построено на основании статистических данных [см.: Гаврилова и др., 1983]](pic/000114.jpg)
Рис. 16. Сопоставление исторической динамики смертности женщин Болгарии и Италии. 1 - общая интенсивность смертности женщин Болгарии в возрасте 40 лет; 2 - общая интенсивность смертности женщин Италии в возрасте 40 лет; 3 - возрастная компонента смертности женщин Болгарии в возрасте 40 лет: 4 - возрастная компонента смертности женщин Италии в возрасте 40 лет. Рассчитано и построено на основании статистических данных [см.: Гаврилова и др., 1983]
Рассмотрим другой пример. В начале XX в. смертность женщин в Болгарии была намного выше, чем в Италии (рис. 16), и также не было никаких оснований считать, что эта ситуация должна измениться. Однако расчет биологической (возрастной) компоненты смертности показал, что между Болгарией и Италией нет существенных различий по величине данной компоненты у женщин. Следовательно, можно было сделать прогноз, что по мере уменьшения социально обусловленной (фоновой) компоненты смертности должна произойти историческая конвергенция смертности - различия в смертности женщин Болгарии и Италии должны существенно уменьшиться. Данное предсказание полностью подтвердилось (рис. 16).
![Рис. 17. Сопоставление исторической динамики смертности женщин Финляндии и мужчин Дании. 1 - общая интенсивность смертности женщин Финляндии в возрасте 40 лет; 2 - общая интенсивность смертности мужчин Дании в возрасте 40 лет; 3 - возрастная компонента смертности женщин Финляндии в возрасте 40 лет; 4 - возрастная компонента смертности мужчин Дании в возрасте 40 лет. Рассчитано и построено на основании статистических данных [см.: Гаврилова и др., 1983]](pic/000115.jpg)
Рис. 17. Сопоставление исторической динамики смертности женщин Финляндии и мужчин Дании. 1 - общая интенсивность смертности женщин Финляндии в возрасте 40 лет; 2 - общая интенсивность смертности мужчин Дании в возрасте 40 лет; 3 - возрастная компонента смертности женщин Финляндии в возрасте 40 лет; 4 - возрастная компонента смертности мужчин Дании в возрасте 40 лет. Рассчитано и построено на основании статистических данных [см.: Гаврилова и др., 1983]
Третий пример состоит в сравнении смертности людей разных полов в разных странах. В начале века смертность женщин Финляндии намного превышала смертность мужчин Дании (рис. 17), и задача прогнозирования таких различий между столь разными группами населения на первый взгляд казалась безнадежной. Тем не менее расчет биологической (возрастной) компоненты смертности позволяет уверенно прогнозировать историческую инверсию смертности, поскольку возрастная компонента у женщин Финляндии намного меньше, чем у мужчин Дании. Современные данные полностью подтверждают правильность сделанного прогноза.
Таким образом, проверяемая концепция позволяет правильно прогнозировать уровни исторической стабилизации смертности и величину различий между этими уровнями для разных популяций. Следует особо отметить, что для такого прогнозирования не требуется никакой информации о причинах смерти и временных тенденциях смертности, а достаточно располагать лишь данными об общей смертности в виде традиционных таблиц дожития почти столетней давности. Насколько нам известно, в современной демографии и биологии человека нет других методов, позволяющих решать подобную задачу. Все это дает основания считать результаты проведенных расчетов не бессмысленным вычислительным упражнением, а объективной оценкой реально существующих явлений, к тому же полезной для практических целей (новые возможности прогнозирования смертности).
Развиваемый подход позволяет также объяснить и количественно прогнозировать увеличение различий в продолжительности жизни мужчин и женщин. Как известно, разрыв в продолжительности жизни мужчин и женщин в начале века составлял всего один-два года. Теперь же эти различия достигают восьми-десяти лет [Lopez, 1983]. Распространено мнение, что поскольку различия выросли сравнительно недавно, то их причину нужно искать в сегодняшнем дне, т. е. в особенностях современного образа жизни [Lopez, 1983]. Однако полученные нами результаты дают основание сделать прямо противоположный вывод. На рис. 18 приведена историческая динамика смертности мужчин и женщин Дании. Видно, что в начале века смертность женщин была близка к смертности мужчин и даже немного ее превышала. Однако на основании этих же таблиц можно рассчитать, что биологическая компонента смертности у женщин несколько меньше. Эти различия в начале века были просто замаскированы высоким уровнем фоновой смертности. Затем в ходе социального прогресса фоновая компонента смертности приблизилась к нулевому значению, и ранее скрытые биологические различия смогли проявиться в полной мере.
![Рис. 18. Сопоставление исторической динамики смертности женщин и мужчин Дании. 1 - общая интенсивность смертности женщин Дании в возрасте 40 лет; 2 - общая интенсивность смертности мужчин Дании в возрасте 40 лет; 3 - возрастная компонента смертности женщин Дании в возрасте 40 лет; 4 - возрастная компонента смертности мужчин Дании в возрасте 40 лет. Рассчитано и построено на основании статистических данных [см.: Гаврилова и др., 1983]](pic/000116.jpg)
Рис. 18. Сопоставление исторической динамики смертности женщин и мужчин Дании. 1 - общая интенсивность смертности женщин Дании в возрасте 40 лет; 2 - общая интенсивность смертности мужчин Дании в возрасте 40 лет; 3 - возрастная компонента смертности женщин Дании в возрасте 40 лет; 4 - возрастная компонента смертности мужчин Дании в возрасте 40 лет. Рассчитано и построено на основании статистических данных [см.: Гаврилова и др., 1983]
Еще более наглядно эта закономерность прослеживается на примере Италии (рис. 19). В начале века смертность мужчин и женщин в этой стране была практически одинаковой. Однако расчет биологической компоненты смертности выявляет большие различия по этой величине. Поэтому можно предсказать, что с течением времени должна произойти историческая дивергенция смертности мужчин и женщин Италии, причем половые различия в смертности должны превысить аналогичные различия, наблюдаемые в Дании. Чтобы убедиться в справедливости этого прогноза, достаточно сопоставить рис. 18 и 19.
![Рис. 19. Сопоставление исторической динамики смертности женщин и мужчин Италии. 1 - общая интенсивность смертности женщин Италии в возрасте 40 лет; 2 - общая интенсивность смертности мужчин Италии в возрасте 40 лет; 3 - возрастная компонента смертности женщин Италии в возрасте 40 лет; 4 - возрастная компонента смертности мужчин Италии в возрасте 40 лет. Рассчитано и построено на основании статистических данных [см.: Гаврилова и др., 1983]](pic/000117.jpg)
Рис. 19. Сопоставление исторической динамики смертности женщин и мужчин Италии. 1 - общая интенсивность смертности женщин Италии в возрасте 40 лет; 2 - общая интенсивность смертности мужчин Италии в возрасте 40 лет; 3 - возрастная компонента смертности женщин Италии в возрасте 40 лет; 4 - возрастная компонента смертности мужчин Италии в возрасте 40 лет. Рассчитано и построено на основании статистических данных [см.: Гаврилова и др., 1983]
Таким образом, в рамках развиваемого подхода можно не только объяснить рост различий в продолжительности жизни мужчин и женщин, но и количественно прогнозировать величину этих различий в каждой конкретной стране. Следует еще раз подчеркнуть, что для этого достаточно иметь лишь традиционные демографические таблицы продолжительности жизни, относящиеся к началу нашего века. Никакой дополнительной информации о структуре причин смерти или о временных тенденциях смертности для этих прогнозов не требуется.
Приведенные примеры доказывают, что обнаруженные половые и региональные различия в уровнях биологической смертности являются объективной реальностью и могут быть использованы для прогнозирования смертности населения. В частности, оценка биологической компоненты смертности позволяет надежно прогнозировать явления исторической конвергенции, дивергенции и инверсии смертности в сравниваемых человеческих популяциях. И хотя некоторым демографам может показаться спорным вывод о том, что Даже при сходных социальных условиях разные человеческие популяции могут иметь разную продолжительность жизни, этот вывод является вполне естественным с точки зрения популяционной генетики и современной эпидемиологии.
|
ПОИСК:
|
При использовании материалов проекта обязательна установка активной ссылки:
http://gelib.ru/ 'Геронтология и гериатрия'