4.3. Понятие о видовых инвариантах продолжительности жизни
Как видно из предыдущего, обсуждение и проверка гипотезы видового предела оказывается настолько увлекательной проблемой, что можно даже забыть, ради чего проводилось это исследование. Поэтому имеет смысл вернуться к началу данной главы и еще раз кратко сформулировать исходную задачу.
Для выяснения причин межвидовых различий по срокам жизни с помощью строгих количественных методов необходима мера, характеризующая продолжительность жизни особей данного вида. Поскольку существуют значительные индивидуальные различия по срокам жизни даже в пределах одной и той же популяции, то возникает проблема выбора искомой величины. Если к тому же учесть, что распределения продолжительности жизни сильно варьируют от популяции к популяции одного и того же вида, то наше положение оказывается даже хуже, чем у Буриданова осла - ему приходилось выбирать только из двух возможностей, в то время как перед нами целый континуум возможных значений.
Выходом из создавшегося положения могло бы быть выявление возраста, обладающего такими особыми свойствами, которыми никакой другой возраст не обладает. В частности, если бы действительно существовал абсолютный верхний предел долголетия, характерный для особей данного вида, то лучшей характеристики видовой продолжительности жизни нельзя было бы придумать. Однако, как мы уже убедились, видовой предел продолжительности жизни ведет себя как мираж, убегающий от нас всякий раз, когда мы пытаемся к нему приблизиться, изучая смертность долгожителей. Поэтому приходится искать другие подходы к определению видовой продолжительности жизни.
Как известно, поиск намного облегчается, когда ищущий знает, что он хочет найти. Если мы ищем величину, характеризующую продолжительность жизни особей данного вида, то она должна быть единой для всех популяций данного вида, независимо от их генетических особенностей и условий жизни. Иначе говоря, важнейшим свойством видовой продолжительности жизни должна быть, ее инвариантность относительно средовых воздействий и генетических особенностей в пределах одного вида. Именно это желаемое свойство инвариантности видовой продолжительности жизни постоянно выдавалось за действительное и приписывалось абсолютному пределу длительности жизни. Как уже неоднократно отмечалось, о видовом пределе продолжительности жизни человека утверждали, что он "оставался неизменным для всех времен, рас и цивилизаций" [Economos, 1985]. И хотя само по себе это утверждение необоснованно, в нем четко сформулированы те желаемые свойства, которыми должна обладать o видовая продолжительность жизни. Действительно, если бы удалось найти величину, характеризующую длительность жизни человека, единую "для всех времен, рас и цивилизаций", то ее можно было бы назвать видовой продолжительностью жизни, даже если бы этот возраст не обладал никакими другими особыми свойствами, кроме инвариантности. Таким образом, инвариантность - тот важнейший критерий, по которому мы можем найти величину видовой продолжительности жизни.
Один из компромиссных подходов к определению видовой продолжительности жизни состоит в выборе такого возраста, до которого доживает лишь небольшая часть популяции [Дубина, Разумович, 1975]. Так, максимальную продолжительность жизни можно определять не как верхний абсолютный предел, а как возраст, до которого доживает лишь 1, 0,1, 0,01 или 0,001% исходной численности популяции. Полученные таким образом оценки, в отличие от рекордов долголетия, уже не будут неограниченно расти с увеличением размера выборки. Более того, теоретически их величина вообще не должна зависеть от числа наблюдений. Для таких оценок можно определить дисперсию и построить доверительные интервалы. В то же время подобная характеристика должна вполне устраивать сторонников гипотезы видового предела, которые могут использовать данную величину, называя ее "практическим пределом продолжительности жизни". В этом случае, к всеобщему удовлетворению, такую величину можно было бы назвать видовой продолжительностью жизни, если бы она была инвариантной в пределах одного вида. Этого, к сожалению, не наблюдается. Величина максимальной продолжительности жизни не удовлетворяет даже критерию исторической инвариантности - она меняется со временем для населения одной и той же страны. Так, например, для населения США возраст, соответствующий выживанию 0,001% новорожденных, составлял в начале века (1900 г) 104,8 года для мужчин и 105,4 - для женщин. В 1980 г. этот возраст увеличился до 111,4 года для мужчин и 113,9 - для женщин [Social Security Administration, 1982]. Поэтому нет никаких оснований утверждать, что величина максимальной продолжительности жизни была неизменной "для всех времен, рас и цивилизаций", если всего лишь за последние 80 лет, когда можно было хоть что-то проверить, она увеличилась на 6,6 года у мужчин и 8,5 - у женщин. В этом увеличении максимальной продолжительности жизни нет, впрочем, ничего удивительного, поскольку даже снижение детской смертности должно было привести к увеличению рассматриваемого показателя. Таким образом, величины подобного типа (соответствующие возрасту, до которого доживает определенная часть популяции) в принципе не могут быть видовыми характеристиками продолжительности жизни.
Другой путь поиска видовой продолжительности жизни состоит не в использовании вероятности дожития до данного возраста 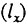 , которая зависит от смертности во всех возрастах, включая детские, а в изучении вероятности смерти в течение года
, которая зависит от смертности во всех возрастах, включая детские, а в изучении вероятности смерти в течение года 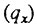 которая отражает смертность только в изучаемой возрастной группе. В научной литературе имеются попытки определения видового предела продолжительности жизни путем экстраполяции возрастной зависимости вероятности смерти до теоретического предельного значения вероятности, равного единице [Depoid, 1973; Халявкин, 1985]. Возраст, соответствующий этому предельному значению вероятности, называется предельной продолжительностью жизни [Халявкин, 1985], хотя на самом деле, с учетом определения вероятности смерти, эта величина'должна быть на один год больше [Depoid, 1973]. Чаще всего подобную экстраполяцию проводят исходя из гипотезы, что вероятность смерти экспоненциально растет с возрастом. Однако, как было показано ранее, такая гипотеза совершенно не согласуется с реальными данными (рис. 26). Если уж вводить гипотезу об экспоненциальном росте, то это следует делать не для вероятности, а для интенсивности смертности. Тогда величина, которую рассчитывают сторонники видового предела, является не абсолютным пределом продолжительности жизни, а просто возрастом, в котором интенсивность смертности близка к единице. Единственная особенность этого возраста состоит в том, что соответствующее ему число доживающих уменьшается в течение года примерно в "e" раз (примерно в 2,7 раза). Ясно, что этой "особенностью" возраста совершенно недостаточно, чтобы считать его видовой характеристикой продолжительности жизни. Однако если бы величина данного возраста оказалась одинаковой для всех популяций человека, то его действительно можно было бы назвать видовой характеристикой продолжительности жизни. В этом случае также был бы возможен разумный компромисс со сторонниками гипотезы предельного возраста, поскольку подобную величину можно было бы назвать "теоретически предельным возрастом" для гипотетической популяции с экспоненциальным ростом вероятности смерти. Однако и этот компромисс оказывается невозможен, поскольку такой "теоретически предельный возраст" не удовлетворяет критерию исторической инвариантности.
которая отражает смертность только в изучаемой возрастной группе. В научной литературе имеются попытки определения видового предела продолжительности жизни путем экстраполяции возрастной зависимости вероятности смерти до теоретического предельного значения вероятности, равного единице [Depoid, 1973; Халявкин, 1985]. Возраст, соответствующий этому предельному значению вероятности, называется предельной продолжительностью жизни [Халявкин, 1985], хотя на самом деле, с учетом определения вероятности смерти, эта величина'должна быть на один год больше [Depoid, 1973]. Чаще всего подобную экстраполяцию проводят исходя из гипотезы, что вероятность смерти экспоненциально растет с возрастом. Однако, как было показано ранее, такая гипотеза совершенно не согласуется с реальными данными (рис. 26). Если уж вводить гипотезу об экспоненциальном росте, то это следует делать не для вероятности, а для интенсивности смертности. Тогда величина, которую рассчитывают сторонники видового предела, является не абсолютным пределом продолжительности жизни, а просто возрастом, в котором интенсивность смертности близка к единице. Единственная особенность этого возраста состоит в том, что соответствующее ему число доживающих уменьшается в течение года примерно в "e" раз (примерно в 2,7 раза). Ясно, что этой "особенностью" возраста совершенно недостаточно, чтобы считать его видовой характеристикой продолжительности жизни. Однако если бы величина данного возраста оказалась одинаковой для всех популяций человека, то его действительно можно было бы назвать видовой характеристикой продолжительности жизни. В этом случае также был бы возможен разумный компромисс со сторонниками гипотезы предельного возраста, поскольку подобную величину можно было бы назвать "теоретически предельным возрастом" для гипотетической популяции с экспоненциальным ростом вероятности смерти. Однако и этот компромисс оказывается невозможен, поскольку такой "теоретически предельный возраст" не удовлетворяет критерию исторической инвариантности.
Так, величина данного предела, рассчитанная совместно для обоих полов на основании статистических данных из Франции, Нидерландов. Швеции и Швейцарии, относящихся к довоенному периоду (30-е годы), составила 110,69 года. Эта же величина, рассчитанная тем же автором для аналогичных данных, относящихся к послевоенному периоду (20-25 лет спустя), оказалась равной 117,28 лет для мужчин и 119,28 - для женщин [Depoid, 1973]. Ф. Депуа, обнаружившая этот феномен увеличения "теоретически предельного возраста", считает его "тем более замечательным, что он, видимо, не имел прецедента в истории" (Ibid). Нам, однако, представляется значительно более примечательным то. что в истории не было ни одного прецедента, когда был бы доказан факт исторической неизменности этого "теоретически предельного возраста". В самом деле, было бы удивительно, если бы данный показатель оставался неизменным, несмотря на снижение смертности пожилых людей.
Следует также отметить, что величина этого теоретически предельного возраста оказывается разной для населения разных стран. Действительно, нетрудно показать, что теоретически предельный возраст (т. е. возраст, когда экстраполированная вероятность смерти равна единице), соответствующий модели экспоненциально роста однолетних значений вероятности смерти, равен:
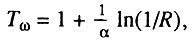
где 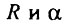 - соответственно предэкспоненциальный множитель и показатель экспоненты в модели экспоненциального роста вероятности смерти. Поскольку, как уже отмечалось, значения вероятности смерти и интенсивности смертности численно близки друг к другу в области малых значений смертности, параметры для модели Гомперца и модели экспоненциального роста однолетних значений вероятности смерти практически совпадают. Поэтому мы можем использовать параметры формулы Гомперца, приведенные в табл. 6, и рассчитать значения теоретически предельного возраста для мужчин и женщин разных стран. Проведя такой расчет, нетрудно убедиться, что значения теоретически предельного возраста неодинаковы для разных популяций человека. Таким образом, данный показатель также не может служить мерой видовой продолжительности жизни, поскольку не удовлетворяет критерию инвариантности.
- соответственно предэкспоненциальный множитель и показатель экспоненты в модели экспоненциального роста вероятности смерти. Поскольку, как уже отмечалось, значения вероятности смерти и интенсивности смертности численно близки друг к другу в области малых значений смертности, параметры для модели Гомперца и модели экспоненциального роста однолетних значений вероятности смерти практически совпадают. Поэтому мы можем использовать параметры формулы Гомперца, приведенные в табл. 6, и рассчитать значения теоретически предельного возраста для мужчин и женщин разных стран. Проведя такой расчет, нетрудно убедиться, что значения теоретически предельного возраста неодинаковы для разных популяций человека. Таким образом, данный показатель также не может служить мерой видовой продолжительности жизни, поскольку не удовлетворяет критерию инвариантности.
Итак, среди предельных характеристик долголетия не удается найти видовой инварианты продолжительности жизни. Поэтому приходится отказаться от поиска компромисса с приверженцами гипотезы видового предела и обратиться к другим характеристикам продолжительности жизни. Это решение тем более оправданно, что экстремальные показатели являются не единственными и далеко не самыми лучшими характеристиками изучаемых распределений. На данное обстоятельство, в частности, обращал внимание Б. Ц. Урланис, отмечая, что вероятность достижения возраста долгожительства ничтожна, "поэтому совершенно неверно рассматривать этот возраст как подлинный человеческий век" [Урланис. 1978, с. 255]. А. А. Зубов и В. И. Козлов, изучая феномен долгожительства, также высказывают мнение, "что рекордное долголетие, вероятно, столь же мало характеризует биологические закономерности старения, как и отдельные рекорды в том или ином виде спорта - средние физические возможности человека" [Зубов, Козлов, 1982, с. 131.
Действительно, если мы хотим определить величину видовой продолжительности жизни и использовать ее в дальнейших исследованиях, то необходимо позаботиться о том, чтобы она была согласована со значениями других видовых характеристик. Поясним сказанное примером. В настоящее время ведутся интенсивные исследования, связывающие величину видовой продолжительности жизни с различными анатомическими, физиологическими, онтогенетическими, биохимическими, а также молекулярно-генетическими характеристиками сравниваемых видов (подробнее см. раздел 5.2). При этом из многих десятков использованных показателей ни один не характеризует крайние рекордные значения изучаемых признаков. Почему же в таком случае мы должны делать исключение для продолжительности жизни? Подобная несообразность, лежащая в основе большинства межвидовых исследований продолжительности жизни, сильно снижает обоснованность полученных результатов, несмотря на безусловную актуальность самой проблемы.
Действительно, хорошо известно, что типичные значения изучаемых признаков характеризуются вовсе не предельными, а средними величинами (средней арифметической, медианой, модой и т. п.). Поэтому если нас интересуют типичные значения продолжительности жизни особей данного вида, то мы должны обратиться именно к средним, а не к максимальным величинам. При этом, однако, снова возникает проблема инвариантности показателей, поскольку все средние величины оказываются неодинаковыми для разных популяций одного и того же вида.
Один из путей решения данной проблемы состоит в расщеплении наблюдаемого распределения продолжительности жизни на составляющие компоненты. При таком подходе можно надеяться на выделение инвариантной компоненты распределения продолжительности жизни, которую можно охарактеризовать соответствующей средней.
Именно таким способом в 1978 г. Б. Ц. Урланисом была определена величина видовой продолжительности жизни человека: 86 лет для мужчин и 88 - для женщин [Урланис. 1978]. Метод определения видовой продолжительности жизни был основан на разложении наблюдаемого распределения числа умерших (dj на три субсовокупности: умерших в результате травм и других неблагоприятных воздействий внешней среды, умерших в результате "нормального социального старения" и, наконец, умерших в сроки, соответствующие "чисто биологическим особенностям человеческого организма" [Урланис. 1978. с. 245-248]. Именно для этой третьей компоненты распределения числа умерших и было определено модальное значение, названное видовой продолжительностью жизни. Следует отметить, что в своих оценках Б. Ц. Урланис исходил из гипотезы нормального распределения биологических сроков жизни, сформулированной еще в 1877 г. В. Лексисом, поэтому значения модальной, медианной и средней (арифметической) продолжительности жизни для третьей субсовокупности умерших совпадали.
Результаты, полученные Урланисом, можно интерпретировать следующим образом: в случае устранения всех неблагоприятных воздействий ("социальных трений", по его определению) распределение людей по срокам жизни будет описываться нормальным законом с модальным значением, равным 86 годам для мужчин и 88 - для женщин. Интересно отметить, что 2 года спустя американский исследователь Дж. Фриз использовал поразительно похожий подход, не зная, по-видимому, о работах Урланиса [Fries, 1980]. Дж. Фриз также исходил из допущения, что распределение по срокам жизни должно следовать нормальному закону, а наблюдаемые отклонения связаны с устранимыми случаями преждевременной смерти. Исходя из такой гипотезы и статистических данных по смертности населения США, Фриз определил, что "при идеальных социальных условиях средний возраст умерших близок к 85 годам" [Fries, 1980, р. 131]. Эта величина, рассчитанная совместно для обоих полов, оказалась весьма близка к оценкам Урланиса, несмотря на приблизительный характер проведенных расчетов [Урланис, 1978. с. 251] и различия в использованных статистических данных.
Разумеется, совпадение результатов, полученных разными исследователями, служит определенным аргументом в их пользу, но не доказывает их справедливости. Более того, имеются некоторые сомнения в обоснованности проведенных расчетов.
Действительно, в своих оценках Урланис и Фриз исходят из гипотезы нормального распределения по срокам жизни "при идеальных социальных условиях". Однако никакого обоснования этой гипотезы авторы не приводят, если не считать высказывания, что "во всех областях жизни господствует какое-то распределение, которое в большинстве случаев подчиняется именно нормальному закону" [Урланис, 1978, с. 240]. Справедливость подобного высказывания представляется довольно спорной, поскольку нет ни одной работы, доказывающей большую распространенность нормального закона по сравнению с другими функциями распределения. Более того, реально наблюдаемые распределения, как правило, асимметричны и характеризуются отличным от нуля эксцессом. Несмотря на эти подчас достоверные отклонения от нормального распределения, анализ данных действительно нередко проводят, принимая за основу гипотезу нормального распределения. Такая практика, однако, отражает не справедливость нормального закона, а лишь стремление исследователей его использовать, поскольку в этом случае можно применять стандартные статистические методы. Следует однако, отметить, что в последние годы все большее число исследователей осознают необходимость перехода к методам непараметрической статистики, не требующим допущения о нормальном распределении изучаемых признаков.
Нет также никаких теоретических оснований предпочитать нормальный закон множеству других законов распределения, известных в теории вероятностей и математической статистике. Более того, можно привести простое соображение, показывающее, что природа в принципе не может быть устроена так, чтобы все изучаемые признаки были распределены по нормальному закону. Например, если мы изучаем распределение частиц по размерам, то для этого параметра можно использовать три меры: линейные размеры (эффективный радиус или диаметр), площадь поверхности и. наконец, их объем (или вес). Нетрудно заметить, что если хотя бы один из изучаемых показателей распределен по нормальному закону, то два других уже в принципе не могут быть распределены по тому же закону, поскольку все три характеристики нелинейно связаны друг с другом. Таким образом, закон распределения зависит даже от выбора единицы измерения признака. Поэтому, даже если какие-то фундаментальные характеристики жизнеспособности организмов и распределены по нормальному закону, из этого вовсе не следует, что распределение наблюдаемых сроков жизни также должно быть нормальным.
Наконец, есть и прямые экспериментальные доказательства непригодности нормального закона распределения для описания вариабельности по срокам жизни. Как известно, распределение людей по срокам жизни характеризуется ярко выраженной левосторонней (отрицательной) асимметрией. Это резкое отклонение наблюдаемых распределений от нормального Б. Ц. Урланис и Дж. Ф. Фриз объясняют существованием преждевременной устранимой смертности, обусловленной "социальными трениями". Справедливость данной гипотезы легко проверить, обратившись к таблицам выживания лабораторных животных, содержащихся в стандартных условиях, где не может быть никакой речи о социальных трениях. Как было показано в гл. 2, распределение продолжительности жизни лабораторных животных резко отличается от нормального, характеризуясь, как правило, той же левосторонней (отрицательной) асимметрией, которая наблюдается в популяциях человека [Семенова, 1983]. Кроме того, при особенно благоприятных условиях содержания распределения продолжительности жизни лабораторных животных имеют ярко выраженный положительный эксцесс, т. е. эти распределения являются значительно более "островершинными" по сравнению с кривой нормального распределения [Там же]. Более подробно проблема распределения продолжительности жизни была разобрана в гл. 2, где показана несостоятельность нормального закона распределения и в то же время достаточно хорошее соответствие реальных данных распределению Гомперца-Мейкема.
Таким образом, приведенные примеры еще раз показывают, что проблема индивидуальных различий по срокам жизни действительно является ключевой в биологии продолжительности жизни. Какие бы задачи мы ни пытались решить, включая определение видовой продолжительности жизни, мы вновь и вновь возвращаемся к проблеме вариабельности организмов по этому признаку.
Действительно, поиск видовых инвариант продолжительности жизни значительно упрощается, если этот процесс разбить на несколько этапов. На первом этапе, когда мы определяем параметры распределения продолжительности жизни для изучаемой популяции, мы практически определяем величины, не зависящие от возраста. Так, например, рассчитав параметры уравнения Гомперца-Мейкема, мы всегда можем определить значения интенсивности смертности в интересующем нас возрасте, однако сами эти параметры от возраста уже не зависят. Затем, сопоставляя эти величины для одной и той же популяции в различные моменты времени, можно выявить параметры, инвариантные относительно времени. В нашем случае таким свойством исторической стабильности обладают возрастная компонента смертности, а также определяющие ее параметры (). Эти характеристики, будучи устойчивы относительно времени, проявляют, однако, значительную региональную изменчивость и зависят от пола. Поэтому следующий естественный шаг исследования состоит в сопоставлении значений этих параметров для разных популяций одного вида в надежде на выявление еще более общих (видовых) инвариант. Такой путь "последовательной экстракции инвариантности" из реальных данных может Оказаться значительно более плодотворным, чем попытки непосредственного определения мировых констант из сырого необработанного материала. В случае же последовательного поэтапного выделения инвариант мы можем обнаружить целую иерархию инвариант разной степени общности, вложенных друг в друга, как в матрешку. При подобном подходе поиск видовых характеристик продолжительности жизни сводится к сопоставлению параметров для разных популяций одного вида. К обсуждению результатов такого исследования мы сейчас и перейдем.
|
ПОИСК:
|
При использовании материалов проекта обязательна установка активной ссылки:
http://gelib.ru/ 'Геронтология и гериатрия'