5.3. Анализ внутривидовых различий по длительности жизни
Анализ внутривидовых различий включает в себя следующие три направления исследований:
- Анализ различий в продолжительности жизни различных линий одного и того же вида. В данном разделе будет приведен конкретный пример подобного подхода.
- Анализ половых различий по срокам жизни. В силу обширности этой проблемы, а также ее специфичности ей будет посвящен специальный раздел (5.4).
- Анализ индивидуальных различий по срокам жизни, поиск предикторов и маркеров индивидуальной продолжительности жизни. Данное направление исследований настолько обширно, что заслуживает специальной монографии. В данной книге эта проблема уже была частично освещена (см. гл. 2).
Хотя во всех перечисленных случаях различия по срокам жизни намного меньше, чем при межвидовых сопоставлениях, однако многие проблемы сопоставимости данных (см. раздел 5.2) при этом практически исчезают. Ниже приведен конкретный пример, иллюстрирующий большие возможности метода внутривидовых сопоставлений.
При изучении механизмов, определяющих продолжительность жизни, собственным признакам организма уделяется несравненно больше внимания, чем аналогичным признакам родителей. Считается очевидным, что связь между признаками родителей и длительностью жизни потомков, даже если она существует, носит не прямой, а опосредованный характер благодаря наследованию тех же признаков потомками. Поэтому при наличии "прямой" информации о собственных признаках организма "косвенная" информация о родительских признаках считается бесполезной для прогнозирования длительности жизни и обычно не используется вообще [Sacher, Duffy, 1979].
Прямо противоположное представление вытекает из предложенной в 1978 г. надежностной концепции продолжительности жизни [Гаврилов. 1978; Гаврилов и др., 1978]. Согласно этой концепции, продолжительность жизни организмов имеет много общего с долговечностью и прочностью материалов: в обоих случаях огромную роль играет исходное распределение дефектов, поскольку именно их цепное лавинообразное размножение определяет динамику последующего разрушения [Гаврилов, 1987]. С этих позиций следует ожидать, что родительские признаки, определяющие качество половых клеток, и особенно материнские признаки, определяющие точность ранних этапов развития, должны иметь гораздо большее влияние на продолжительность жизни потомства, чем те же собственные признаки потомства. Иными словами, надежностная концепция приводит нас к парадоксальному предсказанию: лучшие предикторы продолжительности жизни следует искать не среди собственных признаков организма, а среди родительских признаков.
В данной работе проверено и полностью подтверждено парадоксальное предсказание надежностной концепции продолжительности жизни. Обнаружено неизвестное ранее явление приоритета родительских признаков над собственными признаками при прогнозировании продолжительности жизни. Выявленная закономерность существенно изменяет традиционные представления о механизмах детерминации длительности жизни и открывает новое направление исследований в биологии продолжительности жизни.
Учитывая принципиальное значение обсуждаемой проблемы, особое внимание было уделено качеству исходных данных. С этой целью для анализа были использованы наиболее надежные данные, полученные в Национальной лаборатории Аргонны (США) под руководством известного специалиста Дж. Сэчера. Следует особо подчеркнуть, что эти данные были получены вне связи с проверяемой нами гипотезой и к тому же полностью опубликованы [Sacher, Duffy, 1979], что позволяет любому исследователю независимо проверить корректность полученных нами результатов.
В лаборатории Дж. Сэчера у мышей-самцов с разными генотипами были измерены следующие параметры: средняя продолжительность жизни, вес тела, интенсивность метаболизма по потреблению кислорода на единицу веса в покое (во время сна) и в среднем за сутки. Последние три параметра измерялись дважды: в молодом (6-8 месяцев) и старом (24-34 месяца) возрастах, что соответствовало шести различным переменным. Из 21 исследованного генотипа пять соответствовали следующим инбредным линиям мышей: A/JAnl, BALB/cJAnl, C57BL/6JAnl, C3Hf/JAnl и DBA/IJ. Остальные 16 генотипов соответствовали F1-гибридам, полученным путем скрещивания перечисленных выше инбредных линий в различных комбинациях и направлениях скрещивания.
Анализ данных работы проводился нами следующим образом: средняя продолжительность жизни F1-гибридов рассматривалась как зависимая переменная (продолжительность жизни потомства), значения которой предсказывались на основании 18 других переменных: шести собственных признаков F1-гибридов, шести признаков материнских линий и шести - отцовских. Поскольку число рассматриваемых предикторных переменных (18) превышало число наблюдений (16), был использован метод шаговой регрессии с последовательным включением в уравнение только тех переменных, которые наилучшим образом предсказывают величину продолжительности жизни (программа 2R в известном пакете BMDP).
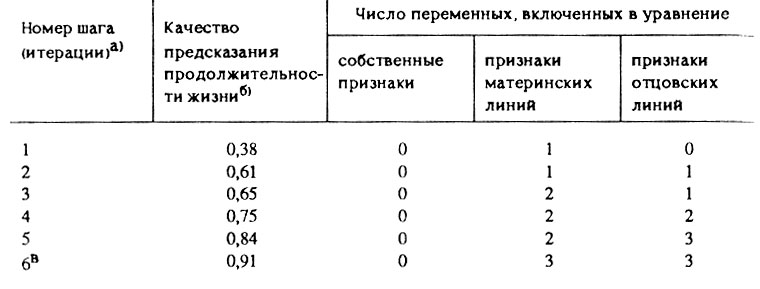
Таблица 15. Приоритет родительских признаков над собственными признаками организмов при прогнозировании продолжительности жизни
а) (На каждом шаге в регрессию включалась одна новая наилучшая предикторная переменная, так что номер шага совпадает с общим числом переменных в уравнении и равен сумме чисел в трех столбцах справа.)
б) (Качество предсказания оценивалось по величине приведенного (скорректированного) коэффициента корреляции, возведенного в квадрат, т. е. с учетом уменьшения числа степеней свободы при увеличении числа переменных.)
в) (После 6-го шага дальнейшее включение новых переменных не приводило к заметному улучшению качества предсказания, а величина R1-критерия для новых переменных не превышала 1.1. Поэтому дальнейшее включение переменных было бессмысленным.)
Результаты проведенного анализа данных превзошли наши самые смелые ожидания. Оказалось, что собственные признаки вообще не позволяют прогнозировать продолжительность жизни, в то время как Родительские позволяют это делать с большой точностью. Самая лучшая" корреляция между продолжительностью жизни и собственным признаком оказалась недостоверной даже при 90% доверительной вероятности (r = -0,39; F(1,14) = 2,51; P>0,1), в то время как четыре родительских признака были связаны с продолжительностью жизни потомков гораздо более тесно. Табл. 15 содержит результаты последовательного включения наилучших предикторов продолжительности жизни в уравнение множественной регрессии. Все 18 переменных были поставлены в равные условия и конкурировали друг с другом за место в уравнении по критерию прогностической ценности. Оказалось, что признаки материнских и отцовских линий включаются в уравнение примерно в равных пропорциях, в то время как собственные признаки не выдерживают конкуренции с родительскими и вообще не включаются в уравнение.
Табл. 16 содержит результаты анализа прогностической ценности четырех различных наборов переменных, шести собственных признаков; шести признаков материнских линий; шести признаков отцовских линий; а также 12 признаков родительских линий (6 материнских и 6 отцовских линий). Оказалось, что все шесть собственных признаков объясняют не более 25% наблюдаемой вариации продолжительности жизни, причем ни один набор переменных с собственными признаками не обеспечивал статистической достоверности регрессии даже при 90%-ном уровне значимости. Для сравнения отметим, что первый же включенный признак материнских линий (минимальная интенсивность потребления кислорода в молодом возрасте) объясняет уже 42% межпопуляционной дисперсии продолжительности жизни потомства с доверительной вероятностью, превышающей 99% (r = -0,65; F(1,14) = 10,14; P<0,01). Если же для прогнозирования используется шесть родительских признаков, то они объясняют не 25 (как в случае собственных признаков), а 95% вариации продолжительности жизни. Таким образом, продолжительность жизни можно точно рассчитывать по значениям родительских признаков, в то время как собственные признаки в этом отношении оказываются практически бесполезными.
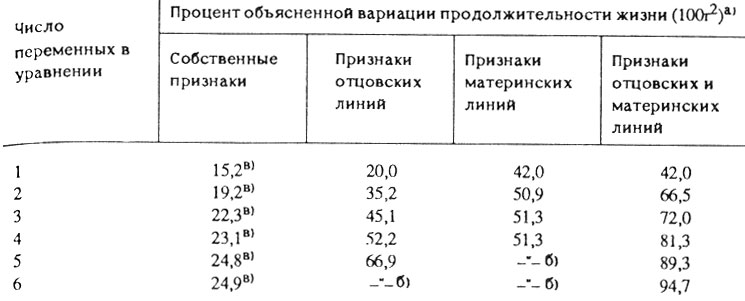
Таблица 16. Сравнение прогностической ценности собственных и родительских признаков
а) (Поскольку на каждом шаге включалась наилучшая предикторная переменная, процент объясняемой дисперсии также является одним из наиболее высоких из возможных при заданном числе и типе переменных.)
б) (Новые переменные не включались в уравнение, поскольку их коэффициент множественной корреляции с другими переменными, уже находящимися в уравнении, был близок к единице и никакого улучшения прогноза ожидать не приходилось. Иначе говоря, толерантность новых переменных оказалась настолько близка к нулю, что дальнейшие расчеты были бессмысленны.)
в) (Регрессия недостоверна даже при 90% доверительной вероятности, что установлено по величине F-критерия.)
Обнаруженное парадоксальное явление приоритета родительских признаков хорошо согласуется с представлением о том, что продолжительность жизни, смертность и заболеваемость во многом определяются событиями, происходящими задолго до рождения и, возможно, даже до образования зиготы (иначе трудно объяснить приоритет отцовских признаков над собственными). Один из возможных механизмов подобной детерминации продолжительности жизни потомства - это накопление повреждений в ДНК половых клеток, вызванных активными побочными продуктами кислородного метаболизма родителей. В последнее время обнаружено, что скорость такого окислительного повреждения ядерной ДНК и особенно митохондриальной ДНК побочными продуктами кислородного метаболизма очень велика [Richter et al., 1988]. В свете этих данных становится понятным, почему из всех исследованных нами признаков именно показатели скорости дыхания родителей оказались наилучшими предикторами продолжительности жизни потомства и почему средняя продолжительность жизни потомства оказывается тем меньше, чем выше скорость дыхания родителей в покое (рис. 33). На рис. 33 приведена зависимость средней продолжительности жизни потомства от скорости дыхания родителей. Скорость дыхания родителей рассчитывалась как полусумма скоростей потребления кислорода молодыми спящими мышами отцовской и материнской линий. Видно, что с увеличением скорости дыхания родителей средняя продолжительность жизни потомства сильно уменьшается (коэффициент корреляции r = -0,81, связь достоверна, P<0,01). Экстраполяция этой зависимости к нулевой скорости дыхания родителей приводит к средней продолжительности жизни потомства, равной 1662 дням. В рамках обсуждаемой гипотезы это означает, что при полной защите половых клеток родителей от окислительной деструкции продолжительности жизни потомства может быть увеличена в среднем в 2 раза! Показательно, что собственная скорость дыхания молодых спящих мышей, в отличие от скорости дыхания родителей, не оказывает влияния на продолжительность жизни (коэффициент корреляции составляет всего -0,1; связь недостоверна, см. рис. 34). Этот факт также хорошо согласуется с обсуждаемой концепцией исключительно ранней детерминации продолжительности жизни.
![Рис. 33. Обратная зависимость между минимальной скоростью дыхания родителей и продолжительностью жизни потомства (гибридных мышей). Скорость дыхания родителей рассчитана как полусумма минимальных скоростей дыхания у отцовской и материнской линий. Коэффициент корреляции r = -0,81. Рассчитано и построено на основании данных, опубликованных в работе [Sacher Duffy 1979]](pic/000164.jpg)
Рис. 33. Обратная зависимость между минимальной скоростью дыхания родителей и продолжительностью жизни потомства (гибридных мышей). Скорость дыхания родителей рассчитана как полусумма минимальных скоростей дыхания у отцовской и материнской линий. Коэффициент корреляции r = -0,81. Рассчитано и построено на основании данных, опубликованных в работе [Sacher Duffy 1979]
Из обсуждаемой концепции следует принципиальная возможность значительного увеличения продолжительности жизни и снижения заболеваемости путем профилактики повреждений половых клеток и нарушений на ранних этапах онтогенеза. В пользу этих представлений свидетельствует также факт увеличения продолжительности жизни потомства при добавлении защищающих от окислительной деструкции антиоксидантов в диету матери [Harman, Eddy, 1979]. По-видимому, этот факт следует рассматривать лишь как первый шаг в очень перспективном направлении "сверхпервичной" профилактики старения и болезней. Что касается фундаментальных исследований биологии продолжительности жизни, то очевидна необходимость развития нового направления исследований по выяснению роли родительских признаков в детерминации продолжительности жизни.
![Рис. 34. Отсутствие выраженной зависимости между продолжительностью жизни и собственной минимальной скоростью дыхания у гибридных мышей. Коэффициент корреляции r = -0,1. Построено на основании данных, опубликованных в работе [Sacher, Duffy, 1979]](pic/000165.jpg)
Рис. 34. Отсутствие выраженной зависимости между продолжительностью жизни и собственной минимальной скоростью дыхания у гибридных мышей. Коэффициент корреляции r = -0,1. Построено на основании данных, опубликованных в работе [Sacher, Duffy, 1979]
|
ПОИСК:
|
При использовании материалов проекта обязательна установка активной ссылки:
http://gelib.ru/ 'Геронтология и гериатрия'