5.4. Анализ половых различий по срокам жизни
Одним из перспективных подходов к выяснению механизмов, определяющих продолжительность жизни организмов, несомненно является изучение причин половых различий по этому признаку. Действительно, о большей продолжительности жизни самок писалось Давно. Еще Дарвин рассматривал меньшую продолжительность жизни самцов как "естественное и конституционное свойство, обусловленное только полом" [Darwin, 1847]. А. Комфорт, проанализировав эту проблему в свете современных данных, также пришел к выводу о большей продолжительности жизни самок у большинства изученных видов, хотя и обратил внимание на ряд исключений [Комфорт, 1967; Comfort, 1979]. На рис. 35 и 36 приведены примеры, подтверждающие большую продолжительность жизни самок по сравнению с самцами.
![Рис. 35. Соотношение между средней продолжительностью жизни мужчин и женщин. Каждая точка соответствует данным по той или иной стране в соответствующее время. Точки, лежащие выше биссектрисы прямого угла (прямая линия), соответствуют случаю, когда продолжительность жизни женщин превышает продолжительность жизни мужчин. Точки, лежащие ниже биссектрисы, соответствуют противоположному случаю. Построено на основании данных, опубликованных в работе [Брук, 1981]](pic/000166.jpg)
Рис. 35. Соотношение между средней продолжительностью жизни мужчин и женщин. Каждая точка соответствует данным по той или иной стране в соответствующее время. Точки, лежащие выше биссектрисы прямого угла (прямая линия), соответствуют случаю, когда продолжительность жизни женщин превышает продолжительность жизни мужчин. Точки, лежащие ниже биссектрисы, соответствуют противоположному случаю. Построено на основании данных, опубликованных в работе [Брук, 1981]
Таким образом, факт большей продолжительности жизни самок как будто бы не вызывает сомнений. В то же время к настоящему времени довольно хорошо изучены генетические, биохимические и физиологические особенности обоих полов. Таким образом, появляется возможность, сопоставляя эти особенности с наблюдаемыми различиями по срокам жизни, высказывать и проверять целый ряд гипотез относительно механизмов, определяющих продолжительность жизни.
![Рис. 36. Соотношение между средней продолжительностью жизни самцов и самок Drosophila mojavensis. График содержит 39 точек, из которых 36 лежат над биссектрисой прямого угла, а 3 - под биссектрисой. Средняя продолжительность жизни самок достоверно чаще превышает среднюю продолжительность жизни самцов (P>0,99). Источник: [Starmer et al., 1977]](pic/000167.jpg)
Рис. 36. Соотношение между средней продолжительностью жизни самцов и самок Drosophila mojavensis. График содержит 39 точек, из которых 36 лежат над биссектрисой прямого угла, а 3 - под биссектрисой. Средняя продолжительность жизни самок достоверно чаще превышает среднюю продолжительность жизни самцов (P>0,99). Источник: [Starmer et al., 1977]
Первая мысль, которая возникает при таком сопоставлении, - это соображение о том, что у женщин имеются две X-хромосомы, а у мужчин - только одна, поэтому генетические дефекты в X-хромосоме в полной мере проявляются у самцов, снижая их жизнеспособность, и практически безвредны для клеток женского организма. Подобное объяснение, предложенное еще в начале века [Комфорт, 1967], оказалось, однако, чрезмерно упрощенным, поскольку выяснилось, что у женщин активна лишь одна X-хромосома [Schneider, 1978]. Тем не менее эта гипотеза сохранилась в несколько модифицированной и своеобразной форме. Так, Д. Харман предполагает, что большая продолжительность жизни самок определяется в первые 48 часов развития зиготы [Harman, 1981]. В этот ранний период функционируют обе X-хромосомы, и, таким образом, наблюдается большая избыточность клеток женского организма по сравнению с клетками мужского. Харман обращает внимание на то, что именно в X-хромосоме находится ген, кодирующий глюкозо-6-фосфатдегидрогеназу - ключевой фермент, участвующий в образовании НАДФ*Н, НАДФ*Н, в свою очередь, имеет существенное значение для поддержания глютатиона в восстановленной форме - мощного ингибитора свободнорадикальных процессов окисления и, кроме того, субстрата глютатионпероксидазы, восстанавливающей перекись водорода до воды и органические гидроперекиси - до соответствующих спиртов. Гипотеза, предложенная Харманом, позволяет также объяснить, почему женщины могут быть менее чувствительны к недостатку селена, входящего в состав глютатионпероксидазы (эта проблема обсуждалась нами ранее в разделе 3.4). В соответствии с гипотезой Хармана можно ожидать, что защитный эффект антиоксидантов, ингибирующих свободнорадикальные процессы, должен сильнее проявляться на менее защищенных представителях мужского пола. Действительно, оказалось, что добавление антиоксидантов в пищу беременных мышей приводит к увеличению продолжительности жизни потомства, причем продолжительность жизни самцов увеличивается при этом в существенно большей степени, чем самок [Harman, Eddy, 1979]. В то же время следует признать, что данная интересная гипотеза нуждается в дальнейшей проверке и более прямых доказательствах.
Важная особенность инактивации одной из X-хромосом на ранних этапах развития женского организма состоит в том, что эта инактивация происходит случайно. В результате женский организм представляет собой удивительную мозаику клеток, в одной половине которых активна отцовская X-хромосома, в другой - материнская [Schneider, 1978]. Такая система клеток в принципе должна быть более устойчивой в случае, когда одна из X-хромосом является дефектной, поскольку другая половина клеток при этом оказывается нормальной. Таким образом, инактивация одной из X-хромосом приводит к тому, что избыточность проявляется не на внутриклеточном уровне, а на уровне клеточных популяций организма. Следует отметить, что наряду с очевидным выигрышем такая избыточность может иметь и некоторые недостатки. Так. например, оказывается, что антигенное разнообразие клеток женского организма значительно выше по сравнению с мужским. Отсюда можно ожидать, что женщины должны чаще страдать от аутоимунных расстройств, что действительно наблюдается [Hazzard, 1986].
Предложенное выше объяснение половых различий по срокам жизни имеет один существенный недостаток. Известно, что у целого ряда организмов гетерогаметными являются не самцы, а самки (например, у бабочек и кур). Тем не менее у этих видов самцы не живут дольше самок, что явно противоречит данной гипотезе [Комфорт, 1967]. Поэтому обсуждаемая гипотеза не может служить общим объяснением большей продолжительности жизни самок.
В нашей стране проблеме половых различий вообще и различий по срокам жизни в частности большое внимание уделено в работах В. А. Геодакяна [Геодакян, 1983а; 19836]. Им разработана оригинальная концепция, объясняющая целый ряд различий между самцами и самками. В. А. Геодакян полагает, что большая продолжительность жизни самок является общебиологическим явлением, и поэтому его объяснение тоже должно быть общебиологическим, основанным на эволюционных принципах. Согласно данной гипотезе, большая вариабельность признаков характерна именно для мужского пола, поэтому основная тяжесть естественного отбора всегда падает на мужскую часть популяции, приводя к ее повышенной смертности. Таким образом, получается, что меньшая продолжительность жизни самцов действительно имеет общебиологическую природу, связанную с тем, что самцы - эволюционный авангард вида, на котором природа проводит свои эксперименты.
Гипотеза Геодакяна на первый взгляд представляется очень привлекательной. При более же внимательном ее анализе можно заметить два недостатка. Первый из них состоит в том. что в данной гипотезе не раскрыт конкретный молекулярно-генетический механизм, приводящий к большей продолжительности жизни самок. Этот недостаток, однако, нельзя считать существенным, поскольку он может быть в принципе устранен в ходе дальнейшего развития и конкретизации данной гипотезы.
Значительно более важный недостаток данной гипотезы то, что она не согласуется с целым рядом фактов. Во-первых, гипотеза Геодакяна предсказывает, что среди долгожителей должно быть больше мужчин. На самом же деле наблюдается прямо противоположная картина [Гаврилов. 1985]. Во-вторых, согласно гипотезе, по мере улучшения условий жизни и снижения смертности различия в продолжительности жизни мужчин и женщин должны стираться, а при особенно благоприятных внешних условиях средняя продолжительность жизни мужчин должна быть даже выше, чем у женщин. Между тем в настоящее время наблюдается прямо противоположный процесс. По мере роста продолжительности жизни растут и различия по этому признаку между мужчинами и женщинами [Бедный. 1979; Lopez, 1983]. Специальное исследование этого процесса показало, что в последние" годы в развитых странах наблюдается ускоренное по сравнению с мужчинами снижение смертности женщин старших возрастов [Gavrilov, Nosov, 1985]. Наконец, в-третьих, следует отметить, что большая продолжительность жизни самок вовсе не является общебиологической закономерностью, несмотря на распространенность такого мнения. Ввиду важности этого момента остановимся на нем более подробно.
Действительно, если обратиться к работам, посвященным проблеме половых различий по срокам жизни, то можно заметить, что вывод о большей продолжительности жизни самок иллюстрируется лишь отдельными примерами [Комфорт, 1967; Геодакян. 1983а; 19836]. Поскольку в научной литературе имеется масса противоречивых данных, то существует опасность, что к тому или иному выводу можно прийти путем вольного или невольного подбора примеров. Для объективного же решения проблемы необходимо анализировать не только те работы, которые посвящены проблеме большей продолжительности жизни самок, но и все остальные работы, где такой цели не ставилось. Если провести такое скрупулезное исследование, то оказывается, что никакой достоверной тенденции к большей продолжительности жизни самок у большинства видов не наблюдается.
![Рис. 37. Соотношение между средней продолжительностью жизни самцов и самок Drosophila melanogaster. График содержит 279 точек, из которых 141 лежит над биссектрисой прямого угла, а 135 - под биссектрисой. Различия в средней продолжительности жизни самцов и самок недостоверны. Источники данных: [Gonzales, 1923; Pearl, Parker, 1924; Steinfeld, 1929; Luers, 1935; Hall, 1969; Van Herrewege, David, 1970; Allemand et al., 1973; Gould, Clark, 1977; Билева и др., 1978; 1981; Bozcuk, 1978; lints et al., 1979; Unlu, Bozcuk, 1979; Giess, 1980; Шахбазов и др., 1980; Некрасова, Шахбазов, 1981]](pic/000168.jpg)
Рис. 37. Соотношение между средней продолжительностью жизни самцов и самок Drosophila melanogaster. График содержит 279 точек, из которых 141 лежит над биссектрисой прямого угла, а 135 - под биссектрисой. Различия в средней продолжительности жизни самцов и самок недостоверны. Источники данных: [Gonzales, 1923; Pearl, Parker, 1924; Steinfeld, 1929; Luers, 1935; Hall, 1969; Van Herrewege, David, 1970; Allemand et al., 1973; Gould, Clark, 1977; Билева и др., 1978; 1981; Bozcuk, 1978; lints et al., 1979; Unlu, Bozcuk, 1979; Giess, 1980; Шахбазов и др., 1980; Некрасова, Шахбазов, 1981]
В этом отношении особенно показателен пример анализа половых различий продолжительности жизни лабораторных дрозофил. Считается твердо установленным, что у этого вида самки живут дольше самцов [Комфорт. 1967]. Действительно, существует много примеров, доказывающих справедливость этого утверждения. Однако если проанализировать подряд всю литературу, где изучалась продолжительность жизни дрозофил, то оказывается, что существует не меньшее число примеров, когда наблюдается прямо противоположная закономерность (рис. 37). Из 279 в 141 случае средняя продолжительность жизни самок превышала среднюю продолжительность жизни самцов, зато в 135 случаях наблюдалась обратная картина. Статистический анализ этих результатов не позволяет обнаружить какой-либо тенденции к большей продолжительности жизни того или иного пола. Полученный нами результат подтверждает вывод Линтса и его соавторов [Lints et U., 1983] об отсутствии тенденции к большей продолжительности жизни самок дрозофил по сравнению с самцами. Этот же вывод был получен при использовании других, более тонких методов анализа данных по продолжительности жизни. Нами были обработаны данные Гонзалеса [Gonzales, 1923] по продолжительности жизни дрозофил линии Old Falmouth, а также мутантов vestigial, arc, speck, black и purple, включая мутантов, несущих различные комбинации этих мутаций [Гаврилов, 1980]. Обработка данных велась путем расчета уравнений множественной линейной регрессии для продолжительности жизни дрозофил в зависимости от наличия той или иной мутации. При наличии изучаемой мутации аргументу приписывалось значение +1. Если же эта мутация отсутствовала, то аргументу приписывалось значение -1. Поскольку всего было изучено пять мутаций и множество их различных комбинаций, то число элементарных переменных было равно пяти. В качестве шестой независимой переменной использовался признак пола, которому приписывалось значение -1 у самцов и +1 у самок. В результате такой обработки данных получилось следующее уравнение множественной регрессии (приводятся лишь наиболее существенные переменные):
Средняя продолжительность жизни дрозофил (дни) = 25,3 дня - 6,0x1 + 3,2x2x3 - 2,3x2 + 2,2x1x4 + ...,* где x1 переменная, отражающая наличие или отсутствие мутации vestigial, x2 - мутации purple, x3 - мутации arc, x4 - переменная, отражающая мужской (x4 = -1) или женский (x4 = +1) пол.
* (Полностью данное уравнение регрессии приведено в работе [Гаврилов. 1980].)
![Рис. 38. Соотношение между средней продолжительностью жизни самцов и самок лабораторных мышей. График содержит 153 точки, из которых 69 лежат над биссектрисой прямого угла, а 83 - под биссектрисой. Различия в средней продолжительности жизни самцов и самок недостоверны. Источники данных: [Lorenz et al., 1955; Neary et al., 1957; Nowell, Cole, 1959; Kohn, Guttman, 1963; Gordon et al., 1966; Sacher, 1966; Hollicroft et al., 1967; Goodrick, 1974; Femandes et al., 1976; Eklund, Bradford, 1977; Holliday, Stevens, 1978; Clapp et aL, 1979; Storer, 1979; Cheney et al., 1980; Thomson et al., 1981]](pic/000169.jpg)
Рис. 38. Соотношение между средней продолжительностью жизни самцов и самок лабораторных мышей. График содержит 153 точки, из которых 69 лежат над биссектрисой прямого угла, а 83 - под биссектрисой. Различия в средней продолжительности жизни самцов и самок недостоверны. Источники данных: [Lorenz et al., 1955; Neary et al., 1957; Nowell, Cole, 1959; Kohn, Guttman, 1963; Gordon et al., 1966; Sacher, 1966; Hollicroft et al., 1967; Goodrick, 1974; Femandes et al., 1976; Eklund, Bradford, 1977; Holliday, Stevens, 1978; Clapp et aL, 1979; Storer, 1979; Cheney et al., 1980; Thomson et al., 1981]
Анализируя эти данные, следует иметь в виду, что комбинации переменных расположены в порядке убывания их влияния. Так, наибольшее влияние на продолжительность жизни дрозофил оказывает мутация vestigial (x1). резко уменьшая среднюю продолжительность жизни дрозофил, что отражено в отрицательном множителе при переменной. Что касается пола (x4), то его влияние оказывается настолько слабым, что он занимает последнее место среди четырех перечисленных переменных. Но особенно важно то, что пол влияет на продолжительность жизни не независимо от мутаций, а в сильном взаимодействии с мутацией vestigial. При наличии этой мутации самки живут дольше самцов, а при ее отсутствии наблюдается прямо противоположная закономерность. Таким образом, в зависимости от генетического фона продолжительность жизни самок может быть как больше, так и меньше, чем у самцов. Этот пример показывает, как опасно делать широкие обобщения по единичным, пусть даже и достоверным случаям большей продолжительности жизни самок.
![Рис. 39. Соотношение между средней продолжительностью жизни самцов и самок сирийского хомячка. График содержит 25 точек, из которых 2 лежат над биссектрисой прямого угла, а 23 - под биссектрисой. Средняя продолжительность жизни самцов достоверно чаще превышает среднюю продолжительность жизни самок (P>0,99). Источник данных: [Kirkman, Yau, 1972]](pic/000170.jpg)
Рис. 39. Соотношение между средней продолжительностью жизни самцов и самок сирийского хомячка. График содержит 25 точек, из которых 2 лежат над биссектрисой прямого угла, а 23 - под биссектрисой. Средняя продолжительность жизни самцов достоверно чаще превышает среднюю продолжительность жизни самок (P>0,99). Источник данных: [Kirkman, Yau, 1972]
На рис. 38-43 приведены другие примеры, показывающие либо отсутствие тенденции к большей продолжительности жизни самок, либо даже наличие прямо противоположной тенденции. Эти примеры показывают, что вывод о большей продолжительности жизни самок как об общебиологической закономерности является недостаточно обоснованным. Поэтому и гипотеза В. А. Геодакяна, исходящая из общебиологических представлений о большей продолжительности жизни самок, оказывается в данном случае неприменимой. Приведенные примеры свидетельствуют о том, что следует искать не общебиологические, а конкретные для каждого вида объяснения половых различий по срокам жизни.
![Рис. 40. Соотношение между средней продолжительностью жизни самцов и самок лабораторных крыс. График содержит 7 точек, из которых 2 лежат над биссектрисой прямого угла, а 4 - под биссектрисой. Различия в средней продолжительности жизни самцов и самок недостоверны. Источник данных: [Hursch etal, 1955]](pic/000171.jpg)
Рис. 40. Соотношение между средней продолжительностью жизни самцов и самок лабораторных крыс. График содержит 7 точек, из которых 2 лежат над биссектрисой прямого угла, а 4 - под биссектрисой. Различия в средней продолжительности жизни самцов и самок недостоверны. Источник данных: [Hursch etal, 1955]
![Рис. 41. Соотношение между средней продолжительностью жизни самцов и самок бабочек-шелкопрядов График содержит 19 точек, из которых 8 лежат над биссектрисой прямого угла, а 11 - под биссектрисой. Различия в средней продолжительности жизни самцов и самок недостоверны как для всех шелкопрядов, так и для каждого вида в отдельности. Источники данных: (Алпатов, Гордеенко, 1932; Osanai, 1978]](pic/000172.jpg)
Рис. 41. Соотношение между средней продолжительностью жизни самцов и самок бабочек-шелкопрядов График содержит 19 точек, из которых 8 лежат над биссектрисой прямого угла, а 11 - под биссектрисой. Различия в средней продолжительности жизни самцов и самок недостоверны как для всех шелкопрядов, так и для каждого вида в отдельности. Источники данных: (Алпатов, Гордеенко, 1932; Osanai, 1978]
В соответствии с этим выводом имеет смысл вернуться к обсуждению половых различий продолжительности жизни человека. В данном случае действительно существует тенденция к большей продолжительности жизни женщин (рис. 35). Разумеется, отчасти половые различия в смертности человека могут быть обусловлены социальными факторами. Однако подобное объяснение не может быть исчерпывающим. Например, повышенная смертность представителей мужского пола наблюдается во всех возрастных группах и даже до рождения, когда о социальных различиях между полами говорить не приходится. По данным ряда исследователей, среди спонтанных выкидышей мужских плодов оказывается значительно больше, чем женских [Byrne, Warburton, 1987; Honore, 1988], причем на ранних стадиях беременности (3 месяца) отношение мужских плодов к женским достигает 3,8 [McMillen, 1979]. Ясно, что для объяснения таких половых различий смертности приходится обращаться к медико-биологическим, а не социокультурным гипотезам. Ранее уже отмечалось, что половые различия в сроках жизни людей связаны с биологической компонентой смертности и по-разному выражены в разных популяциях человека (см. раздел 3.4). Однако даже в пределах одного этого вида механизм половых различий в смертности может быть неодинаков в разных возрастах. Если во взрослых возрастах изменение условий жизни приводит примерно к одинаковому изменению смертности мужчин и женщин (рис. 44), то в младенческом возрасте (до года) мальчики оказываются значительно более чувствительными к изменению внешних условий (рис. 45). Таким образом, различия в смертности двух полов даже чисто феноменологически могут быть связаны с двумя разными факторами - фактором смертности, не зависящим от внешних условий, но зависящим от возраста (что соответствует разности в величине возрастной компоненты смертности мужчин и женщин), и фактором большей чувствительности к повреждающим воздействиям внешней среды. Из приведенных рисунков видно, что один из механизмов половых различий характерен для взрослых, а другой - для младенцев. Таким образом, мы приходим к необходимости дальнейшего расчленения проблемы половых различий уже в пределах одного и того же вида, но для разных возрастных групп.
![Рис. 42. Соотношение между средней продолжительностью жизни самцов и самок Drosophila subobscura График содержит 11 точек, из которых 4 лежат над биссектрисой прямого угла, а 7 - под биссектрисой. Различия в средней продолжительности жизни самцов и самок недостоверны. Источник данных: [Clark, Rockstein, 1964]](pic/000173.jpg)
Рис. 42. Соотношение между средней продолжительностью жизни самцов и самок Drosophila subobscura График содержит 11 точек, из которых 4 лежат над биссектрисой прямого угла, а 7 - под биссектрисой. Различия в средней продолжительности жизни самцов и самок недостоверны. Источник данных: [Clark, Rockstein, 1964]
![Рис. 43. Соотношение между средней продолжительностью жизни самцов и самок жука-кожееда Trogoderma versicolor. График содержит 9 точек, из которых 5 лежат над биссектрисой прямого угла, а 4 - под биссектрисой. Различия в средней продолжительности жизни самцов и самок недостоверны. Источник данных. [Davis, 1945]](pic/000174.jpg)
Рис. 43. Соотношение между средней продолжительностью жизни самцов и самок жука-кожееда Trogoderma versicolor. График содержит 9 точек, из которых 5 лежат над биссектрисой прямого угла, а 4 - под биссектрисой. Различия в средней продолжительности жизни самцов и самок недостоверны. Источник данных. [Davis, 1945]
Если провести дальнейший анализ различий в продолжительности жизни обоих полов у человека и исследовать смертность от отдельных причин, то обнаруживаются новые неожиданные закономерности. В результате исследований, проведенных нами совместно с В. Г. Семеновой, было обнаружено, что причины смерти сильно различаются по степени взаимосвязанности показателей смертности мужчин и женщин. В качестве примера в табл. 17 приведены значения коэффициента половой согласованности (коэффициента корреляции между показателями смертности мужчин и женщин) для некоторых из 84 изученных причин смерти.
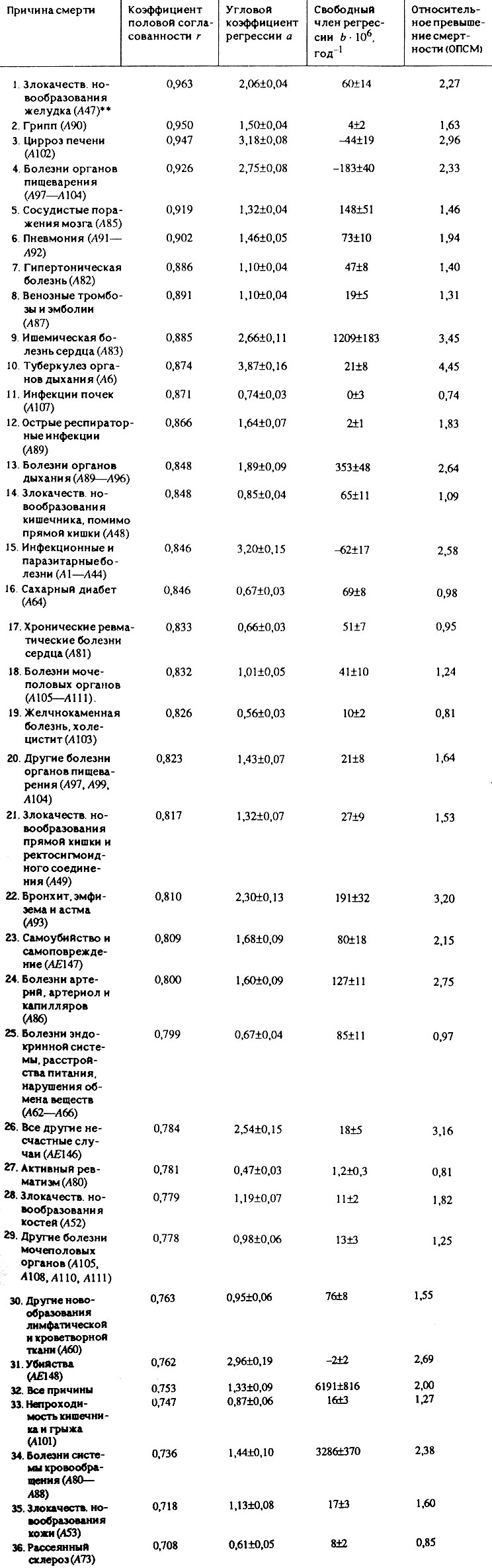
Таблица 17. Характеристики причин смерти, согласованных по полу*
* (Для расчета были использованы статистические данные ВОЗ по 26 разным странам мира за период с 1965 по 1987 г. в возрастной группе 55-64 года (всего 179 реализаций).)
** (В скобках приведены коды причин смерти в соответствии с Перечнем А Международной статистической классификации болезней, травм и причин смерти (VIII пересмотр).)
Можно заметить, что для ряда. болезней (рак желудка, цирроз печени, инсульт, гипертоническая болезнь, грипп и др.) значения смертности мужчин и женщин настолько тесно согласованы между собой, что смертность представителей одного пола можно надежно определить по величине смертности другого пола (рис. 46-50). Однако для других болезней (рак легкого, пищевода, полости рта, гортани) коэффициент половой согласованности оказался поразительно мал - менее 0,5 (рис. 51-53).
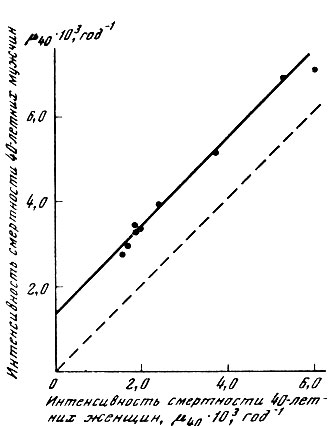
Рис. 44. Линейное соотношение между интенсивностью смертности мужчин и женщин в историческом аспекте. Использованы данные таблиц продолжительности жизни белого населения США за 1920-1929, 1929-1939, 1939-1941, 1949-1951, 1959-1961, 1964, 1970, 1975, 1977 гг.
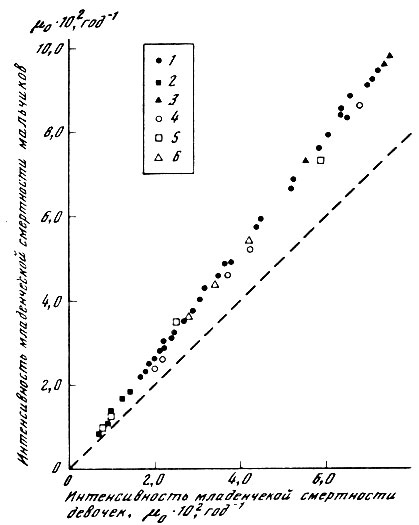
Рис. 45. Линейное соотношение между интенсивностью младенческой смертности мальчиков и девочек. 1 - Франция. Данные за 1931-1937, 1947-1965, 1967, 1969 гг.: 2 - Швеция. Данные за 1956-1960, 1961-1965, 1966-1970, 1971-1975, 1975-1979 гг.; 3 - Дания. Данные за 1921-1925, 1926-1930, 1936-1940 гг.; 4 - США (цветное население). Данные за 1939-1941, 1949-1951, 1964, 1975, 1977 гг.; 5 - Норвегия. Данные за 1911-1920, 1946-1950, 1971-1975, 1976-1980 гг.; 6 - Швейцария. Данные за 1933-1937, 1941-1950, 1948-1953 гг.
Для выяснения механизма обнаруженных различий по степени половой согласованности нами был проведен факторный анализ смертности от причины, согласованной (рак желудка) и несогласованной (рак легкого) по полу. Выяснилось, что географическая и временная изменчивость смертности от рака желудка определяется всего одним фактором, общим для мужчин и женщин. В то же время смертность от рака легкого определялась уже двумя факторами, причем один из них определял смертность мужчин, а другой - женщин (табл. 18). Таким образом, было установлено, что согласованность смертности по полу обеспечивается действием общих (полонеспецифических) факторов, а несогласованность по полу - действием полоспецифических факторов.

Таблица 18. Факторная структура смертности от злокачественных новообразований желудка, а также трахеи, бронхов и легкого*
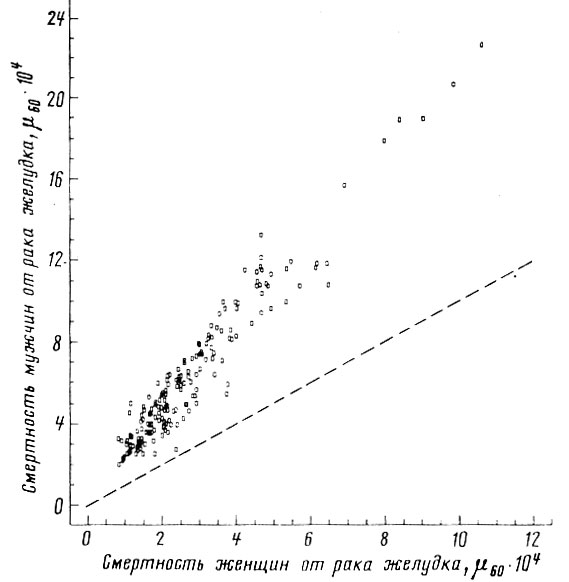
Рис. 46. Корреляция между смертностью мужчин и женщин в возрасте 55-64 лет от злокачественных новообразований желудка. Коэффициент корреляции
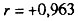 . Пунктиром проведена прямая, соответствующая одинаковой смертности мужчин и женщин. Построено на основании данных для 26 стран (включает 179 реализаций)
. Пунктиром проведена прямая, соответствующая одинаковой смертности мужчин и женщин. Построено на основании данных для 26 стран (включает 179 реализаций)Если повышенная смертность мужчин обусловлена действием дополнительных факторов риска, специфических для мужского пола, то наибольший половой диморфизм смертности должен наблюдаться для причин, не согласованных по полу. Действительно, оказалось, что наибольшее относительное превышение смертности мужчин* (ОПСМ) наблюдается именно для тех причин, которые имеют низкие значения коэффициента половой согласованности. Так, для возрастной группы 55-64 года наибольшее ОПСМ наблюдалось в случае смертности от рака гортани (ОПСМ = 11,8; r = 0,128); несчастных случаев, вызванных огнестрельным оружием (ОПСМ = 11,4; r = 0,395); несчастных случаев производственного характера (ОПСМ = 9,5; r = 0,463); несчастных случаев, связанных с немототранспортными средствами (ОПСМ = 6,2; r = 0,593); рака легкого (ОПСМ = 6,1; r = 0,432).
* (Относительное превышение смертности мужчин рассчитывалось как отношение средней смертности мужчин (усредненной по исследованным странам) к средней смертности женщин в возрастной группе 55-64 года.)
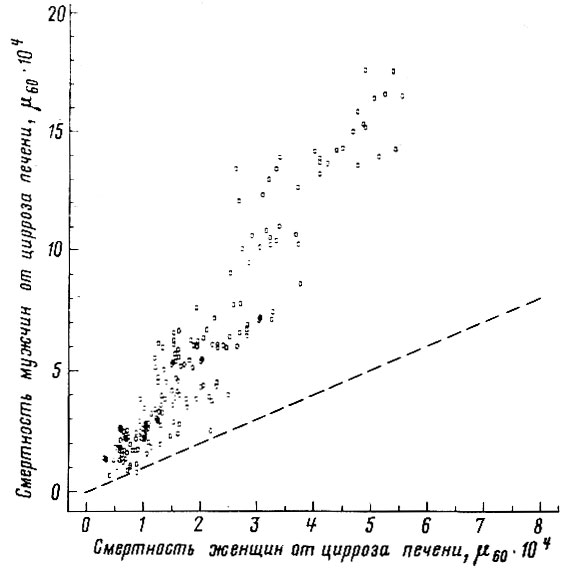
Рис. 47. Корреляция между смертностью мужчин и женщин в возрасте 55-64 лет от цирроза печени. Коэффициент корреляции
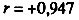 . Пунктиром проведена прямая, соответствующая одинаковой смертности мужчин и женщин. Построено на основании данных для 26 стран (включает 179 реализаций)
. Пунктиром проведена прямая, соответствующая одинаковой смертности мужчин и женщин. Построено на основании данных для 26 стран (включает 179 реализаций)Вместе с тем оказалось, что вклад не согласованных по полу причин смерти в наблюдаемый половой диморфизм общей смертности парадоксально мал и не превышает вклада согласованных по полу причин смерти. Одно из возможных объяснений обнаруженного парадокса состоит в том, что половые различия могут быть обусловлены такими полоспецифическими факторами, которые, мало варьируя в пространстве и времени, не нарушают половой согласованности. В этом случае, если рассчитать коэффициенты уравнения линейной регрессии смертности мужчин по смертности женщин, следует ожидать, что половые различия для согласованных по полу причин смерти должны определяться величиной свободного члена регрессии, отражающего сдвиг в значениях смертности, обусловленный действием инвариантных полоспецифических факторов. Действительно, оказалось, что свободный член регрессии вносит существенный вклад в величину полового диморфизма смертности от согласованной по полу причины. Например, для ишемической болезни сердца этот вклад достигает 33.7% от наблюдаемых половых различий. Вместе с тем очевидно, что свободный член регрессии не является основным источником половых различий в смертности от причин, согласованных по полу.
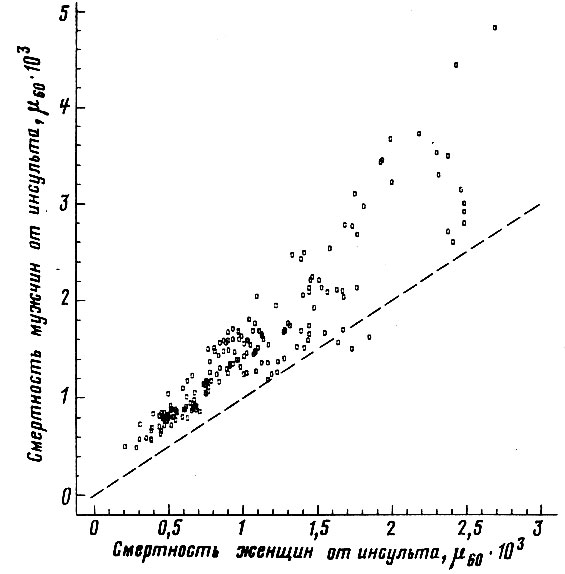
Рис. 48. Корреляция между смертностью мужчин и женщин в возрасте 55-64 лет от сосудистых поражений мозга. Коэффициент корреляции
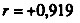 . Пунктиром проведена прямая, соответствующая одинаковой смертности мужчин и женщин. Построено на основании данных для 26 стран (включает 179 реализаций)
. Пунктиром проведена прямая, соответствующая одинаковой смертности мужчин и женщин. Построено на основании данных для 26 стран (включает 179 реализаций)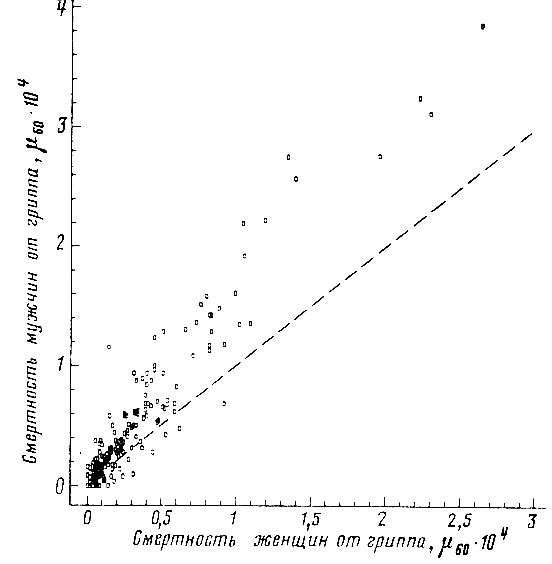
Рис. 49. Корреляция между смертностью мужчин и женщин в возрасте 55-64 лет от гриппа. Коэффициент корреляции r = +0,950. Пунктиром проведена прямая, соответствующая одинаковой смертности мужчин и женщин. Построено на основании данных для 26 стран (включает 179 реализаций)
Из табл. 17 видно, что угловой коэффициент линейной регрессии для согласованных по полу причин смерти значительно больше единицы, т. е. мужская субпопуляция оказывается более чувствительной к действию тех же факторов, которые действуют и на женскую субпопуляцию. Как показали наши исследования, именно эта закономерность вносит основной вклад в половой диморфизм смертности от причин, согласованных по полу, и существенный вклад в половой диморфизм общей смертности. Обнаруженная закономерность может быть использована в дальнейших исследованиях биологии человека, а также для прогнозирования половых различий в смертности людей.
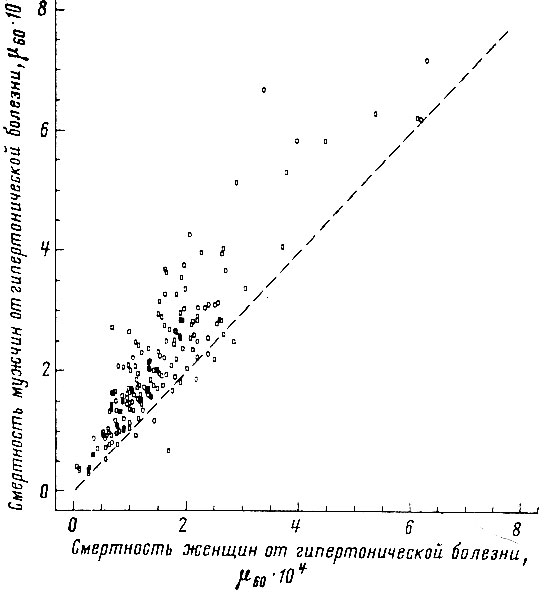
Рис. 50. Корреляция между смертностью мужчин и женщин в возрасте 55-64 лет от гипертонической болезни. Коэффициент корреляции r = +0,886. Пунктиром проведена прямая, соответствующая одинаковой смертности мужчин и женщин. Построено на основании данных для 26 стран (включает 179 реализаций)
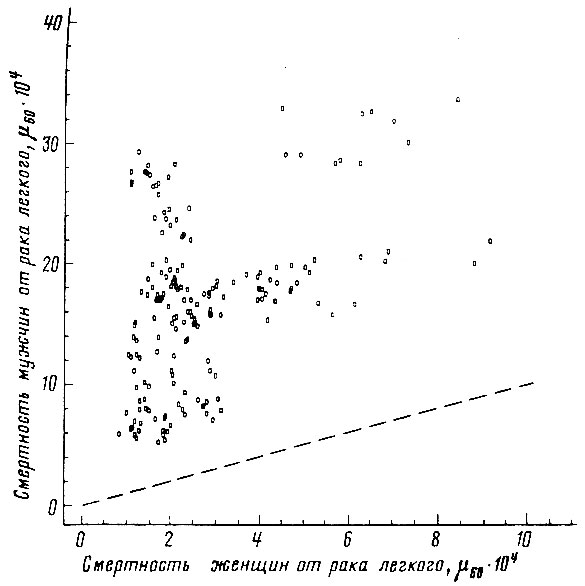
Рис. 51. Соотношение между смертностью мужчин и женщин в возрасте 55-64 лет от злокачественных новообразований трахеи, бронхов и легкого. Коэффициент корреляции r = +0,432. Пунктиром проведена прямая, соответствующая одинаковой смертности мужчин и женщин. Построено на основании данных для 26 стран (включает 179 реализаций)
Подводя итоги попыткам выяснить механизмы, определяющие продолжительность жизни путем изучения половых различий по этому признаку, можно сказать следующее. Надежды, связанные с поиском общебиологических механизмов этих различий, по-видимому, не оправдались, поскольку, вопреки широко распространенному мнению, сама по себе большая продолжительность жизни самок не является общебиологической закономерностью. Даже в пределах одного биологического вида (человек) есть основания для раздельного анализа этой проблемы у разных возрастных групп. Естественно, что такие ограничения значительно снижают оптимизм в исследовании данного вопроса, однако они способствуют лучшему пониманию реальных процессов, приводящих к половым различиям по срокам жизни.
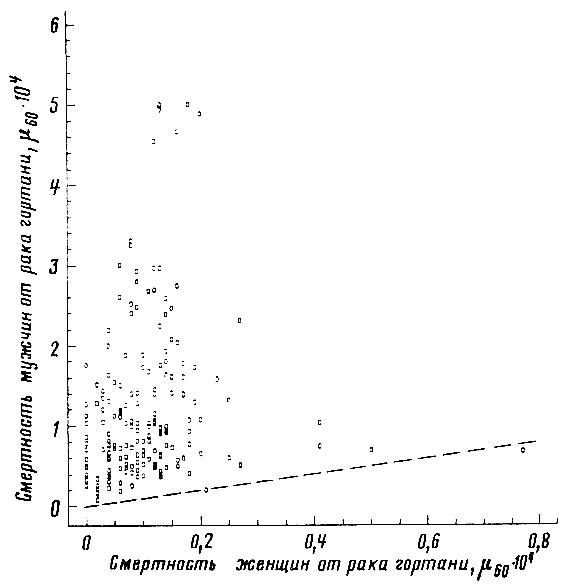
Рис. 52. Соотношение между смертностью мужчин и женщин в возрасте 55-64 лет от злокачественных новообразований гортани. Коэффициент корреляции r = +0,128. Пунктиром проведена прямая, соответствующая одинаковой смертности мужчин и женщин. Построено на основании данных для 26 стран (включает 179 реализаций)
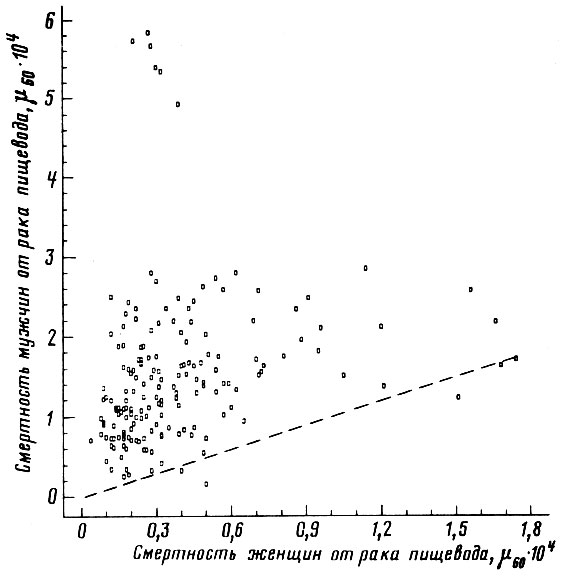
Рис. 53. Соотношение между смертностью мужчин и женщин в возрасте 55-64 лет от злокачественных новообразований пищевода. Коэффициент корреляции r = +0,208. Пунктиром проведена прямая, соответствующая одинаковой смертности мужчин и женщин. Построено на основании данных для 26 стран (включает 179 реализаций)
|
ПОИСК:
|
При использовании материалов проекта обязательна установка активной ссылки:
http://gelib.ru/ 'Геронтология и гериатрия'