6.7. Модель гетерогенной популяции
В двух предыдущих разделах была рассмотрена ситуация, когда варьирование по степени резервирования блоков происходило внутри каждого организма, а сами организмы считались исходно идентичными друг другу и одинаковыми по риску смерти. Последнее допущение может быть оправдано в двух случаях.
Во-первых, однородность популяции может наблюдаться тогда, когда существует жесткая генетическая программа, определяющая исходную степень резервирования каждого блока (органа) в организме. Иными словами, возможно, что механизм генерации вариабельности блоков вовсе не является случайным, а изложенные в двух предыдущих разделах модели оказываются применимы лишь потому, что генетически запрограммированное распределение блоков по степени их резервирования можно аппроксимировать распределением Пуассона или биномиальным распределением. Отметим, что в данном случае популяция организмов исходно однородна и в отношении риска отказов конкретных блоков (однородность в отношении смертности от отдельных причин).
Во-вторых, организмы могут оказаться практически одинаковыми по распределению числа блоков с разной степенью резервирования и при случайном механизме формирования резерва, если только число m незаменимых блоков очень велико. В результате популяция будет практически однородной по риску гибели организмов, несмотря на то что в отношении риска отказов конкретных блоков (соответствующих отдельным причинам смерти) популяция может быть исключительно гетерогенна. Если же число незаменимых блоков в организме сравнительно невелико или если нас интересуют закономерности смертности в результате отказа конкретных блоков (моделирование смертности от отдельных причин), то учет гетерогенности популяции становится настоятельной необходимостью.
В данном разделе будет показано, что учет гетерогенности популяции позволяет объяснить все основные закономерности смертности даже в простейшем случае, когда организм состоит всего из одного блока с n элементами.
Пусть q - вероятность элемента быть исходно работоспособным. Тогда вероятность встретить организм с i исходно работоспособными элементами из общего числа n элементов задается законом биномиального распределения:
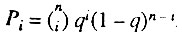
Вероятность организма с i исходно работоспособными параллельно соединенными элементами умереть до возраста x равна:
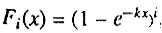
где k - постоянная интенсивность отказа элементов.
Функция распределения времени жизни F(x) для смеси организмов c = 0, 1, 2, ... , n исходно работоспособными элементами определяется следующим образом:
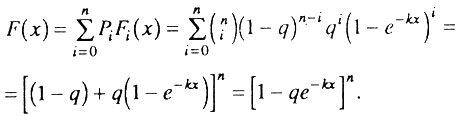
Интенсивность смертности в данном случае равна
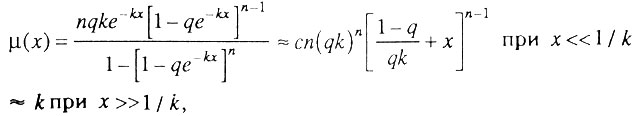
где 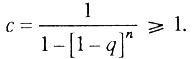
Таким образом, интенсивность смертности в гетерогенной популяции организмов сначала растет с возрастом в соответствии с биномиальным законом смертности, а затем асимптотически стремится к верхнему пределу, равному k.
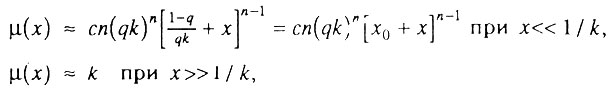
где 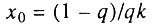 - параметр, который мы предлагаем называть начальным виртуальным возрастом популяции. Действительно, этот параметр имеет размерность времени и соответствует возрасту, в течение которого исходно однородная популяция накопила бы столько дефектных организмов, сколько реальная популяция уже имеет в начальный момент времени (при x = 0). В частности, когда q = 1, т. е. когда все элементы в каждом организме исходно работоспособны, начальный виртуальный возраст популяции равен нулю и интенсивность смертности растет с возрастом по степенному закону (закону Вейбулла), что соответствует случаю, разобранному в разд. 6.5. Когда же популяция исходно неоднородна (q<1), мы приходим к уже упоминавшемуся биномиальному закону смертности. Таким образом, предложенная здесь модель гетерогенной популяции также может служить теоретическим обоснованием биномиального закона смертности.
- параметр, который мы предлагаем называть начальным виртуальным возрастом популяции. Действительно, этот параметр имеет размерность времени и соответствует возрасту, в течение которого исходно однородная популяция накопила бы столько дефектных организмов, сколько реальная популяция уже имеет в начальный момент времени (при x = 0). В частности, когда q = 1, т. е. когда все элементы в каждом организме исходно работоспособны, начальный виртуальный возраст популяции равен нулю и интенсивность смертности растет с возрастом по степенному закону (закону Вейбулла), что соответствует случаю, разобранному в разд. 6.5. Когда же популяция исходно неоднородна (q<1), мы приходим к уже упоминавшемуся биномиальному закону смертности. Таким образом, предложенная здесь модель гетерогенной популяции также может служить теоретическим обоснованием биномиального закона смертности.
В случае, когда 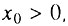 всегда существует начальный период времени, такой, что
всегда существует начальный период времени, такой, что 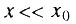 и справедливо следующее приближение биномиального закона:
и справедливо следующее приближение биномиального закона:
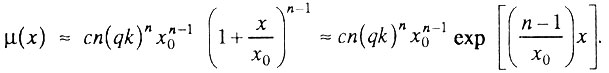
Следовательно, при любом значении q<1 всегда существует начальный период времени x, когда интенсивность смертности экспоненциально растет с возрастом:
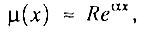
где 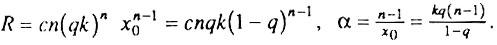
Итак, если популяция исходно гетерогенна (q<1), то интенсивность смертности в начальный период времени экспоненциально растет с возрастом (по закону Гомперца).
Нетрудно заметить поразительное сходство формул и комментариев в данном и предыдущем разделах книги. Это сходство обусловлено тем, что две принципиально разные модели приводят тем не менее к практически совпадающим конечным выводам. Следует, однако, подчеркнуть, что речь идет лишь о внешнем сходстве поведения двух принципиально разных конкурирующих моделей, каждая из которых заслуживает тщательного анализа, несмотря на поразительное внешнее сходство в поведении своего конкурента. Поэтому не следует удивляться, если и дальнейший анализ модели гетерогенной популяции будет похож на анализ модели последовательно соединенных блоков с варьирующей степенью резервирования, рассмотренной в предыдущем разделе книги.
Модель гетерогенной популяции позволяет объяснить не только экспоненциальное увеличение интенсивности смертности с возрастом, но и компенсационный эффект смертности, описанный в разделе 4.5. Действительно, согласно введенным обозначениям:
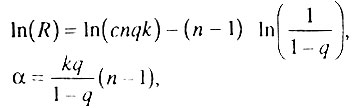
т. е величины 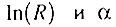 параметрически связаны через величину (n - 1), что позволяет представить
параметрически связаны через величину (n - 1), что позволяет представить 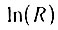 как функцию α:
как функцию α:
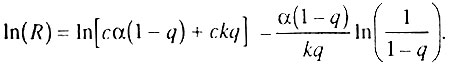
Таким образом, компенсационный эффект смертности наблюдается тогда, когда различия в смертности обусловлены межпопуляционными различиями по числу элементов в организме (n), а другие параметры, включая скорость старения (скорость необратимой гибели элементов k), практически одинаковы для всех сопоставляемых популяций одного вида. Нетрудно заметить сходство подобного объяснения компенсационного эффекта смертности с объяснениями, вытекающими из предыдущих моделей, описанных в разд. 6.5 и 6.6.
Как уже отмечалось (см. разд. 6.6), продолжительность периода экспоненциального роста интенсивности смертности зависит от величины q. В общем случае возрастная динамика смертности популяции на участке возрастов 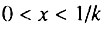 в зависимости от величины q может быть описана тремя сценариями:
в зависимости от величины q может быть описана тремя сценариями:
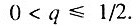 Данный случай соответствует ситуации, когда исходно работоспособно менее половины общего числа элементов. В этом случае
Данный случай соответствует ситуации, когда исходно работоспособно менее половины общего числа элементов. В этом случае 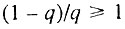 и, следовательно,
и, следовательно, 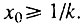 Поэтому во всем интервале, когда
Поэтому во всем интервале, когда 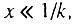 всегда выполняется и условие
всегда выполняется и условие 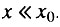 В этом случае интенсивность смертности во всем рассматриваемом интервале экспоненциально растет с возрастом.
В этом случае интенсивность смертности во всем рассматриваемом интервале экспоненциально растет с возрастом.
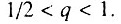 Данный случай соответствует ситуации, когда исходно работоспособно более половины всех элементов. В этом случае
Данный случай соответствует ситуации, когда исходно работоспособно более половины всех элементов. В этом случае 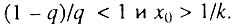 В данной ситуации возрастная динамика смертности в исследуемом возрастном интервале
В данной ситуации возрастная динамика смертности в исследуемом возрастном интервале 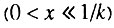 состоит из двух стадий:
состоит из двух стадий:
- первая стадия начального периода, когда
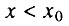 и, следовательно, биномиальный закон смертности сводится к закону Гомперца;
и, следовательно, биномиальный закон смертности сводится к закону Гомперца;
- вторая стадия начального периода, когда
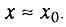 и применим только биномиальный закон смертности в полном виде без приближений.
и применим только биномиальный закон смертности в полном виде без приближений.
- первая стадия начального периода, когда
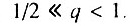 Данный случай соответствует ситуации, когда исходно дефектна лишь очень небольшая часть элементов. В этом случае
Данный случай соответствует ситуации, когда исходно дефектна лишь очень небольшая часть элементов. В этом случае 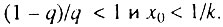 Возрастная динамика смертности в этом случае состоит из трех стадий:
Возрастная динамика смертности в этом случае состоит из трех стадий:
- первая стадия начального периода, когда
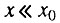 и биномиальный закон смертности сводится к закону Гомперца;
и биномиальный закон смертности сводится к закону Гомперца;
- вторая стадия начального периода, когда
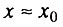 и когда применим только биномиальный закон смертности.
и когда применим только биномиальный закон смертности.
- третья стадия начального периода, когда и
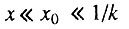 биномиальный закон смертности сводится к степенному закону роста интенсивности смертности (закону Вейбулла).
биномиальный закон смертности сводится к степенному закону роста интенсивности смертности (закону Вейбулла).
- первая стадия начального периода, когда
По мере того как q стремится к единице, продолжительность первой стадии начального периода с экспоненциальным ростом интенсивности смертности стремительно уменьшается, а третьей стадии - стремительно растет. В пределе, в случае исходно однородной популяции (q = 1), мы приходим к закону Вейбулла, справедливому на всем возрастном интервале 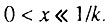
Таким образом, модель гетерогенной популяции позволяет объяснить основные закономерности смертности организмов: экспоненциальный рост интенсивности смертности в начальный период с последующим замедлением темпов ее роста, а также компенсационный эффект смертности. Кроме того, обсуждаемая модель
позволяет выяснить условия, при которых наблюдается не экспоненциальный, а степенной закон роста интенсивности смертности (закон Вейбулла). Наконец, предложенная модель позволяет установить что два на первый взгляд взаимоисключающих закона - Гомперца и Вейбулла,- на самом деле являются лишь частными случаями одного, более общего биномиального закона смертности, поручившего в данной модели строгое теоретическое обоснование.
Согласно модели гетерогенной популяции, судьба неработоспособных элементов и их гибель совершенно не влияют на выводы модели. Поэтому модель будет справедлива и в том случае, если все неработоспособные элементы уже погибнут к моменту формирования взрослого организма, и он будет состоять только из работоспособных элементов (клеток). Важно, что при этом все равно останется след в виде биномиального распределения организмов в популяции по числу исходно работоспособных элементов. Собственно говоря, в этом и состоит суть модели, а рассуждения об исходно дефектных элементах - лишь одно из возможных обоснований причин вариабельности организмов по степени их резервирования. Поэтому предложенная модель может быть названа также моделью популяции организмов с различной степенью резервирования.
С учетом сделанных замечаний основной вывод модели может быть переформулирован следующим образом: Если отдельные особи популяции различаются по степени их резервирования, то интенсивность смертности в начальный период времени экспоненциально растет с возрастом (по закону Гомперца). Это утверждение справедливо при любой форме биномиального распределения организмов: с отрицательной (левосторонней) асимметрией, полностью симметричной (нормальный закон распределения), и с положительной (правосторонней) асимметрией. Единственное влияние формы распределения состоит в том, что при отрицательной асимметрии распределения экспоненциальный рост интенсивности смертности может продолжаться недолго, а в случае симметричного распределения и распределения с положительной асимметрией период экспоненциального роста интенсивности смертности значительно более продолжителен и охватывает весь начальный период, когда 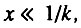
Итак, модель гетерогенной популяции приводит в принципе к тем же выводам, что и ранее рассмотренная модель последовательно соединенных блоков с варьирующей степенью резервирования. Между тем речь идет о принципиально разных моделях: если в одной индивидуальный риск гибели одинаков для всех организмов и экспоненциально растет с возрастом, то в другой исходно существует n субпопуляций живых организмов с различным риском гибели, растущим с возрастом по степенному, а не экспоненциальному закону.
Совпадение конечных выводов, полученных из двух принципиально разных моделей означает, что на основании установленных закономерностей смертности нельзя однозначно выбрать истинный механизм возрастного разрушения организмов и необходимы дальнейшие исследования для проверки конкурирующих моделей. Это обстоятельство, конечно, не может радовать, зато есть два других момента, вселяющих оптимизм. Во-первых, оказывается, что разные модели приводят к практически совпадающим трактовкам некоторых закономерностей смертности. Например, компенсационный эффект смертности, согласно любой из моделей, возможен лишь при видовом постоянстве скорости необратимых возрастных изменений. Такая трактовка компенсационного эффекта смертности содержится не только в трех моделях, последовательно разобранных в разделах 6.5, 6.6 и 6.7 данной книги, но также и в других моделях [Strehler, Mildvan, 1960; Гаврилов, 1978; Гаврилов и др., 1978]. Поэтому надежная содержательная интерпретация ряда закономерностей смертности возможна и при наличии многих конкурирующих теорий, поскольку плюрализм моделей не исключает их "консенсуса" по ряду проблем. Во-вторых, если самые разные модели приводят к одним и тем же формулам, например к биномиальному закону смертности, это затрудняет лишь возможность интерпретации результатов для теоретика, но существенно облегчает работу экспериментатора. В самом деле, при обработке данных предпочтительнее использовать именно ту формулу, за которой стоит не одна, а целое семейство принципиально разных моделей, охватывающих широкий спектр возможных ситуаций.
Изложенная в данном разделе модель гетерогенной популяции позволяет объяснить все основные закономерности смертности даже в простейшем случае, когда организм состоит из одного блока с n параллельно соединенными элементами. Обобщение модели на случай m последовательно соединенных блоков в каждом организме не представляет сложностей, если блоки независимы друг от друга в отношении их надежности. В этом случае естественно ожидать, что интенсивность смертности гетерогенной популяции равна просто сумме интенсивностей отказов всех блоков. В частном случае, когда параметры, определяющие надежность блоков (n, q и k), одинаковы для всех блоков, интенсивность смертности равна произведению числа блоков (m) на интенсивность отказа одного блока. В результате мы приходим к формулам, в точности совпадающим с формулами, полученными в разделе 6.6 для модели однородной популяции.
|
ПОИСК:
|
При использовании материалов проекта обязательна установка активной ссылки:
http://gelib.ru/ 'Геронтология и гериатрия'