Д. Миелограмма у стариков
* (Исследования были проведены совместно с П. Муйкэ.)
Цитоз крови стариков еще недостаточно хорошо изучен.
Советская школа внесла значительный вклад в дело изучения данной проблемы (Моначевич, Соловьев, Бродин, Инушкин и Грагуерова).
Мы исследовали костный мозг у 20 стариков в возрасте от 65 до 92 лет. При этом мы не старались установить какую-либо формулу клеточного состава костного мозга, а стремились дать интерпретацию различным клеточным элементам крови в определенный возрастной период, а именно, в старости.
Для получения костного мозга производилась пункция грудины.
Все время мы стремились избегать неправильных интерпретаций, стараясь сделать скорее общий обзор, чем установить количественное определение форменных элементов. При определении формулы мы учитывали все ядерные элементы за исключением мегакариоцитов и клеток, находящихся в состоянии кариокинеза.
По данным Шиллинга, тип миелограммы стариков соответствует зрелому костному мозгу, а с точки зрения реактивности можно говорить об умеренной реакции по классификации Барта.
Приводим ниже среднее значение миелограммы:

Гранулоэритробластический коэффициент Понтони, варьирующий, по мнению некоторых авторов, в пределах 2-3, в наших наблюдениях изменялся в пределах 2,4-3,6.
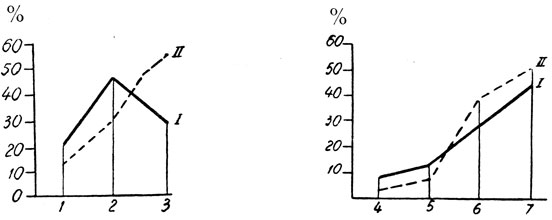
Рис. 103. - Кривые созревания эритроцитобластического и гранулоцитобластического рядов: 1 - Базофильный эритробласт; 2 - Полихроматофильный эритробласт; 3 - Ортохроматофильный эритробласт; 4 - миелобласт; 5 - Промиелоцит; 6 - Миелоцит; 7 - Метамиелоцит: I - Кривая созревания у взрослых; II - Кривая созревания у стариков
Мы отметили кривые созревания для эритроцитобластического и гранулоцитобластического рядов в отдельности (рис. 103).
Кривая созревания гранулоцитов с сдвигом вправо несколько ускорена. Что касается кривой созревания эритроцитов, то в 2% случаев отмечается запаздывание, а в остальных случаях - ускорение.
Юные клетки (миелобласты, промиелоциты) обладают характером зрелых элементов, т. е. их величина уменьшается до 15 µ с незначительным числом ядрышек; в этих клетках наблюдается незначительная азурофильная мелкая зернистость. Большинство клеток обладает хорошо выраженной архоплазмой. Отношение между поверхностью ядрышка и поверхностью ядра тоже снижено, а потенциальность деления весьма незначительна.
Цитоплазма носит слегка базофильный характер. Миелоциты включаются в группу А, по классификации Доана, с незначительным числом зерен, диаметр которых едва достигает 12 µ.
Метамиелоциты как сегментированные, так и палочкоядерные обладают незначительным диаметром, достигающим максимум 10 µ.
Костный мозг обладает слабовыраженной эритропоэтической функцией. В большинстве случаев форменными элементами являются ацидофильные нормобласты, достигающие в диаметре 7-8 µ с небольшим сильно окрашенным ядром.
Эритроциты в диаметре достигают 7,5-8 µ, причем, наблюдаются неравномерные по величине элементы (анизоцитоз).
Ретикулоциты составляют 15-18%, в большинстве случаев включаясь в группу IV, по классификации Гейльмейера, являясь таким образом "старыми" ретикулоцитами.
Мегакариоцитарный ряд весьма незначителен, составляет лишь 1-2 ‰, с диаметром, доходящим до 30-40 µ, со старческим ядром и с отсутствием тромбоцитопоэтической функции. Развитие ядра соответствует эволюционной стадии цитоплазмы.
Гистиоцитарный ряд представлен редкими элементами (2-3%) размером в 30 µ с ярко окрашенным ядром, с 2-3 ядрышками и зернистостью.
Оксидазная формула (определенная по методу Шульце) для гранулоцитобластического ряда в 50% случаев оказалась положительной, в 30% случаев - слабо положительной и в остальных случаях - очень слабо положительной или даже отрицательной.
Зернистость в 60% случаев мелкая, "порошкообразная", а в 40% случаев крупная.
Реакция Фелгена на тимонуклеиновую кислоту оказалась положительной для ядер эритробластического ряда и слабо положительной для ядер гранулоцитобластического ряда.
Митотический коэффициент равнялся 1-2% для красной крови; для белой крови коэффициент не был установлен.
Калиевая реакция оказалась резко положительной для протоплазмы эритробластов, главным образом, в отношении клеточной периферии.
Развитие ядра соответствует эволюционной стадии цитоплазмы. Не было отмечено никаких отклонений от нормального цитологического развития. Вакуоли были обнаружены как в цитоплазме, так и в ядре с диаметром 1-2,5 µ довольно часто, а именно, в 45% случаев.
В некоторых случаях была обнаружена слабо положительная реакция на липиды (Судан III).
Гистологические срезы показали наличие бедной костномозговой ткани, однако со значительным количеством жиров.
Кусочки костного мозга грудины выращивались in vitro в присутствии тимонуклеиновой и рибонуклеиновой кислот; наблюдения продолжались 24-48 часов. Для гранулоцитобластического ряда получалась кривая с сдвигом влево, причем промиелоциты достигали в диаметре 27 µ, миелоциты - 18 µ., а взрослые элементы 12-13,5 µ; отмечалась обильная зернистость, рассеянная по всей протоплазме. Ядрышки варьировали в размерах от 2 до 3 µ. Уже не отмечалось ядерно-цитоплазматического эволюционного синхронизма, а наоборот, проявлялся не ярко выраженный асинхронизм; развитие хроматина происходило более быстро, чем ядрышка. Все вышеуказанные эволюционные изменения имели тенденцию к образованию юных клеток. В красной крови преобладали полихроматофильные нормобласты. Красные кровяные тельца подвергались изменениям и становились микроцитами.
Оксидазная реакция оказалась резко положительной для гранулоцитобластического ряда в 60% случаев. Реакция Фелгена была также резко положительной. Все вышеуказанные изменения более выражены в культурах с присутствием рибонуклеиновой кислоты.
На фоне этого в общем нормального состояния следует все же отметить наблюдавшееся более или менее выраженное нарушение равновесия кроветворной функции. Цитологическое развитие крови проходило через те же стадии, что и организм в процессе старения.
|
ПОИСК:
|
При использовании материалов проекта обязательна установка активной ссылки:
http://gelib.ru/ 'Геронтология и гериатрия'