X. Исследования по экспериментальной иликибиологии
Чтобы лучше уяснить природу явлений в зависимости от возраста, мы считали необходимым прибегнуть не только к методу прямых наблюдений но и к их экспериментальному изучению.
В первую очередь мы занялись исследованием экспериментального процесса старения.
Главной и руководящей идеей при этом являлось следующее положение:
Процесс старения вызывается расстройством неустойчивого и относительного равновесия, существующего между явлениями ассимиляции и диссимиляции, с тенденцией к повышению последних, независимо от того, идет ли речь о действительном усилении последних явлений или о выраженном снижении первых.
В ряде опытов* мы исходили из идеи, что удаление щитовидной железы замедляет в значительной степени явления ассимиляции, а также биохимические явления, психосоматические функции в целом и что с многих точек зрения атиреоз приближается, если вообще не сливается, с одним из видов процесса старения.
* (Опыты были проведены совместно с М. Питиш и А. Ботез.)
С другой стороны, максимум энергии организма, стимуляции его функций в значительной степени наслаивается на пубертатный период, на эпоху юношества и молодости, а значительное число экспериментальных исследований показало, что парентеральное введение мужских половых гормонов в состоянии (до известной степени) возвратить жизненную силу одряхлевшему организму.
Кастрация - также относительный вид процесса старения; это не означает, что она обязательно ведет к укорачиванию длительности жизни; кастрация уменьшает количество жизни, так как жизненный цикл кастратов развивается обыкновенно в замедленном темпе.
Согласно некоторым концепциям, возможно все же, что недостаточность половых желез, главным образом мужских половых желез, представляет собой, по-видимому, удлинение детского периода (сохранение вилочковой железы и т. д.).
Как бы то ни было, мы предприняли исследования в смысле комбинации указанных двух оперативных вмешательств; другими словами, мы удаляли у крыс обоего пола, с одной стороны, половые железы, а с другой - щитовидную железу.
В действительности же, вместе с щитовидной железой удалялась и околощитовидная ткань, так что по крайней мере у некоторых из наших животных мы произвели тройную эндокринную недостаточность.
Особенно интересным фактом в этих опытах является то, что животные вели себя различно во время опыта, а именно, феномены задержки развития были гораздо более выраженными у животных мужского пола.
Однако и у последних кастрация, без других вмешательств, не произвела особо значительных изменений. Удаление щитовидной железы вызвало общеизвестные симптомы. Все же различия по отношению к контрольной группе даже в этих случаях не представляли особого значения.
Комбинирование вышеуказанных двух хирургических операций вызывало весьма выраженную задержку в развитии, старческий вид животных и сокращение длительности жизни.
Опыты были проведены примерно на 300 животных, из которых большинство - крысы, а остальные - морские свинки и кролики.
Животные подвергались наблюдению в течение 2-5 месяцев, что соответствует максимальному выживанию животных с удаленными щитовидной и половыми железами.
Еженедельно мы определяли вес тела, соматические изменения и поведение животного. Так как в первую очередь мы старались установить длительность жизни, то лишь незначительная часть животных была забита и по этой причине нам не удалось во всех случаях взять для исследования органы подопытных животных.
В последнем ряде исследований мы попытались применить лечение тирео-гонадной недостаточности, пользуясь при этом препаратами, отличавшимися от средств, необходимых для корригирования гипофункции эндокринных желез. Так, например, в этой группе мы вводили эпифизгормон и витамин Е, т. е. препараты, давшие хорошие результаты при лечении процесса старения у человека.
Опыты можно сгруппировать следующим образом:
1. Крысы обоего пола в возрасте от 10 до 12 дней с весом тела от 10 до 15 г. Эти животные были разбиты на 4 группы. В первой группе находились крысы с удаленными половыми железами, во второй группе - животные с удаленной щитовидной железой. В третью группу мы поместили животных с одновременно удаленными половыми и щитовидной железами, а последняя группа являлась контрольной. В этой серии выжили лишь животные, подвергнутые удалению половых желез, а также крысы, входившие в контрольную группу.
2. Крысы обоего пола с весом тела примерно 50 г и с приблизительным возрастом в 2 месяца. Эти животные были также разбиты на 3 группы по 20 особей в каждой. Животные первой группы подверглись удалению половых желез. У животных второй группы была удалена щитовидная железа, а у животных третьей группы была произведена комбинированная операция удаления как половых, так и щитовидной желез. Из каждой группы выбиралось несколько особей для контроля. Все животные находились под наблюдением в течение 5 месяцев.
3. Крысы обоего пола с весом тела от 100 до 120 г и с приблизительным возрастом от 5 до 5 1/2 месяцев. Здесь также были образованы 3 подопытные группы по 20 оперированных животных в каждой, как и в предыдущих случаях. 20 животных были оставлены в качестве контрольной группы. Животные находились под наблюдением лишь в течение 3 месяцев.
4. Крысы-самцы с весом тела от 100 до 120 г и с точным возрастом 5 месяцев; 25 из указанных животных подверглись комбинированной операции удаления половых и щитовидной желез, а 5 животных были оставлены в качестве контрольной группы.
5. Крысы-самцы с весом тела от 70 до 80 г, с точным возрастом 4 месяцев. Из всего количества животных 30 крыс подверглись комбинированной хирургической операции, а 10 остались в качестве контрольной группы. Часть оперированных животных принимала парентеральным путем, начиная со дня последней операции, эпифизгормон и витамин Е в дозе 1 мл ежедневно в течение 3 месяцев.
6. Морские свинки-самцы с весом тела от 270 до 450 г, без точно установленного возраста, подвергшиеся операции удаления щитовидной железы или половых желез, а также комбинированной операции удаления как щитовидной, так и половых желез.
7. Кролики-самцы с весом тела от 900 до 1200 г, без точно установленного возраста, разбитые на подобные же группы.
В первом опыте, произведенном на крысах в возрасте 12 дней, отмечалось, что животные с удаленной щитовидной железой или подвергнутые комбинированной операции удаления как щитовидной, так и половых желез, погибли через несколько дней после хирургического вмешательства, а выжили лишь животные, подвергнутые удалению только половых желез, а также животные контрольной группы. Следует отметить, что у животных, подвергнутых удалению половых желез в этом возрасте, наблюдались серьезные расстройства в повышении веса тела (рис. 155 и 156). Так, например, в то время как крысы-самцы контрольной группы прибавили в весе через два месяца в среднем на 450%, а самки - на 525%, животные, подвергнутые удалению половых желез, прибавили в весе тела на 338% и соответственно на 367%. Указанные изменения в весе тела вследствие удаления половых желез в очень раннем возрасте заставили нас произвести специальные исследования в этом направлении.
Во второй серии опытов, произведенных на крысах в возрасте 2 месяцев, различия были более четкими в зависимости от пола (рис. 157 и 158). Так, по отношению к контрольной группе, у самцов с удаленными половыми железами наблюдалось легкое повышение веса тела, в то время как вес самок слегка снизился сравнительно с интактными животными того же пола. Не отмечалось симптомов постарения или значительных изменений в поведении. Волос самцов был более тонким и мягким, приближаясь к волосу самок. В течение опытов не погибло ни одно из животных указанной группы.
У крыс, подвергнутых удалению щитовидной железы, отмечалось значительное замедление в повышении веса и роста тела. В течение 5 месяцев указанные животные прибавили в весе тела на 55% сравнительно с исходным весом, в то время как крысы контрольной группы прибавили в весе тела на 236%. Из этих животных пало 3 в промежутке между первым и вторым месяцем после хирургического вмешательства. У животных отмечалось состояние апатии, зябкость и снижение аппетита по сравнению с контрольной группой. Шерсть была более грубой, жесткой, однако не отмечалось выпадения волос. Недостаточность щитовидной железы оказывала менее значительное влияние на крыс-самок той же группы. Повышение веса тела через 5 месяцев равнялось 120% по сравнению с 216% у контрольных животных. Погибло только одно животное на второй месяц после операции. Самки были менее апатичны по сравнению с оперированными самцами. И в этих случаях шерсть была более грубой и жесткой.
У крыс, подвергнутых удалению щитовидной и половых желез, наблюдались исключительно выраженные симптомы постарения. У самцов повышение веса тела почти полностью прекращалось. Так, например, через 5 месяцев после операции повышение веса тела равнялось лишь 11% по сравнению с исходным весом. Среди указанных животных 3 пало в течение 4-го и 5-го месяцев после операции. Начиная с первого же месяца отмечалось выпадение шерсти, главным образом, на спине и животе, причем через 3 месяца у подопытных животных почти полностью отсутствовала шерсть в указанных областях. Оставшаяся шерсть была грубой и жесткой, а кожа сморщенной, жесткой и покрытой чешуйками (рис. 159). Зубы длинные, желтые и шатающиеся, как у старых животных. У подопытных животных наблюдалось состояние апатии, зябкости, они старались забиться в угол клетки. Совершенно исчезла живость, характерная для этих животных. Отсутствовала также агрессивность. Аппетит также исчез у подопытных животных. Крысы-самки этой группы подвергались менее значительному влиянию в результате комбинированной двойной операции, приводящей к недостаточности щитовидной и половых желез. Самки прибавили в весе тела на 77% по сравнению с исходным весом. Шерсть этих животных была более грубой и жесткой, но не отмечалось выпадения волоса. В общих чертах животные особенно не отличались от самок, подвергнутых лишь простой операции удаления щитовидной железы.
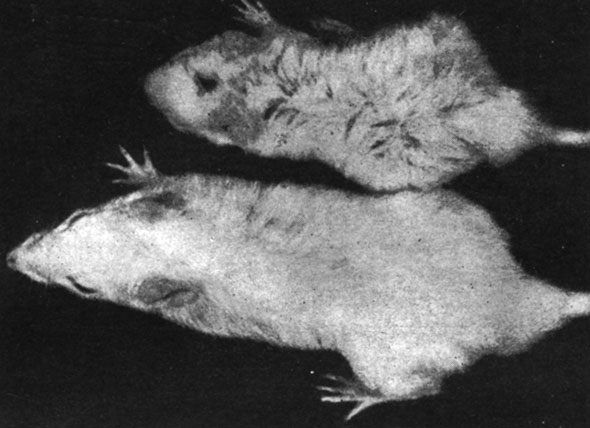
Рис. 155. - Крыса-самец, подвергшаяся удалению половых желез, и контрольное животное
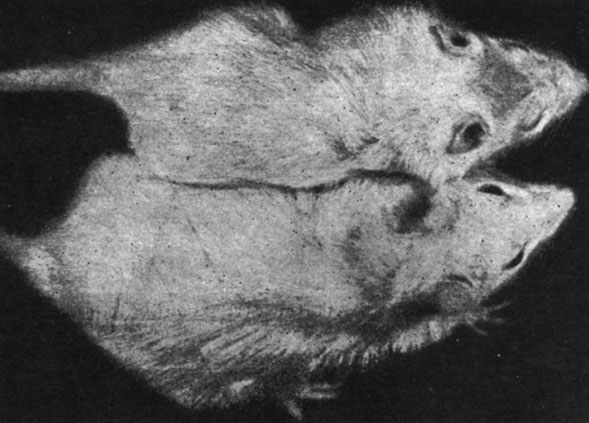
Рис. 156. - Крыса-самка, подвергшаяся удалению половых желез, и контрольное животное
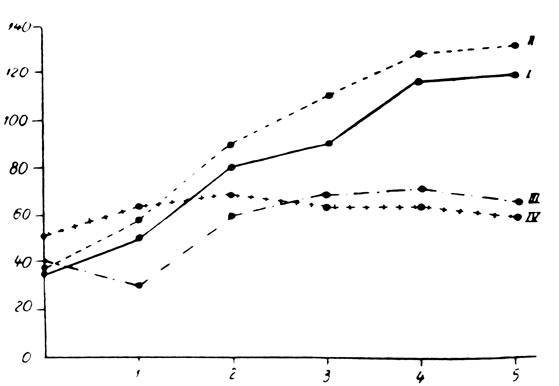
Рис. 157. - Динамика веса тела крыс-самцов, подвергнутых удалению щитовидной и половых желез, по сравнению с контрольной группой: I - Контроль; II - Животные, подвергнутые удалению половых желез; III - Животные, лишенные щитовидной железы; IV - Животные, подвергнутые комбинированной операции удаления щитовидной и половых желез
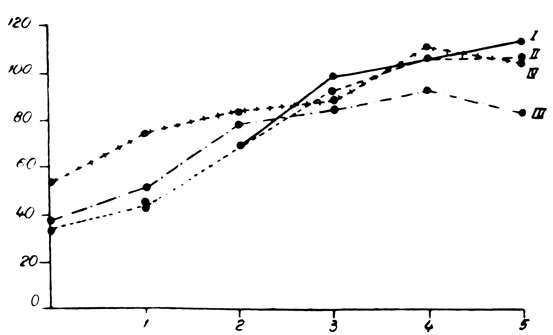
Рис. 158. - Динамика веса тела крыс-самок, подвергнутых удалению щитовидной и половых желез, по сравнению с контрольной группой: I - Контроль; II - Животные, подвергнутые удалению половых желез; III - Животные, лишенные щитовидной железы; IV - Животные, подвергнутые комбинированной операции удаления щитовидной и половых желез
Оставшиеся в живых животные были забиты через 5 месяцев. Гистологическое исследование эндокринных желез подтвердило результаты, полученные посредством наблюдения животных, причем отмеченные сдвиги различались не только в связи с одножелезистой или двужелезистой недостаточностью, но зависели также от пола животных. Так, например, в нижнем мозговом придатке отмечалось повышение числа базофильных клеток, более выраженное у самок, чем у самцов, что относилось как к животным, подвергавшимся удалению половых желез, так и к животным, лишенным щитовидной железы. Указанная базофильная реакция была также значительно более выраженной при двужелезистой недостаточности, чем при одножелезистой, что вновь подтверждает наличие соотношения синергидного типа между гонадотропными и тиреотропными гормонами. Что касается коркового вещества надпочечников, то следует отметить, что у самок наблюдалась реакция гипертрофического типа как при удалении половых желез, так и при удалении щитовидной железы, с умеренными симптомами жирового перерождения лишь у животных с тиреогонадной недостаточностью. У самцов реакция дегенеративного типа сопровождалась иногда обильными кровоизлияниями: в пучковом слое у некоторых животных с тиреоидной недостаточностью и в более выраженной степени у животных с тиреогонадной недостаточностью.

Рис. 159. - Крысы-самцы, подвергнутые удалению щитовидной и половых желез, и контрольное животное (слева)
Третья серия опытов имела целью изучение тех же эндокринных гипофункций у животных более старшего возраста (приблизительно 5-5 1/2 месяцев). И в этих группах поведение подопытных животных отличалось в зависимости от пола, а в общем были получены подобные же изменения, но с менее выраженной интенсивностью. Этот факт, по-видимому, можно объяснить тем, что животные находились под наблюдением сравнительно непродолжительное время, так как начиная с первого месяца после хирургического вмешательства наблюдалась значительная смертность как у животных, подвергнутых удалению щитовидной железы, так и у животных, подвергнутых комбинированной операции удаления щитовидной и половых желез. Так, среди первых животных погибли 2 самца и 1 самка в первый же месяц , 3 самца и 1 самка в течение 2-го месяца и 2 самки в течение 3-го месяца, причем этот срок пережили лишь 4 животных. Среди животных, подвергавшихся комбинированной двойной хирургической операции, погибли 3 самца и 1 самка в первый же месяц, 4 самца и 2 самки в течение 2-го месяца и 1 самец и 3 самки в течение 3-го месяца. Возможно, что смертность обусловливалась отчасти временем года, в котором были проведены опыты, в том смысле, что подопытные животные весьма неустойчивы к холоду.
В четвертой серии опытов мы задались целью проследить лишь эффект двойной хирургической операции на 30 крысах-самцах в возрасте 5 месяцев. У оперированных животных наблюдались те же явления, что и в первой группе. Следует отметить в особенности выраженные изменения зубов: резцы с измененной трофикой, с неправильным ростом, желтовато-серой окраской и резкой расшатываемостью. Указанные изменения следует поставить в связь и с известной степенью околощитовидной недостаточности, хотя, как было указано выше, видимые околощитовидные железы не удалялись. Вес тела животных снизился за время наблюдения на 46% по сравнению с исходным, в то время как в контрольной группе он повысился на 27%. Более 50% животных погибло в первые же 2 месяца, остальные были забиты с целью изучения гистологических изменений.
В пятой серии опытов мы поставили перед собой задачу проследить эффект эпифизгормона и витамина Е на животных, подвергнутых двойной железистой недостаточности (рис. 160). Группа состояла из 40 животных, из которых 10 служили в качестве контрольных, а 30 были подвергнуты двойному хирургическому вмешательству. Среди последних некоторым крысам вводился парентерально эпифизгормон, другим - витамин Е. Вес тела и рост животных, подвергнутых удалению щитовидной и половых желез, без последующего лечения, ничем не отличались от предыдущих групп. То же самое можно сказать о поведенческих реакциях и кожных изменениях, а также и о трофических расстройствах зубов.
У животных, подвергнутых удалению щитовидной и половых желез с последующим лечением витамином Е, снижение веса тела было менее значительным в первый месяц лечения, а затем кривая приобрела вид, встречающийся обыкновенно у нелеченных животных. После этого срока смертность была подобной, отмечавшейся в группе животных после 3 месяцев лечения, причем выжила только одна крыса.
У животных, подвергнутых лечению эпифизгормоном, наблюдались весьма незначительные снижения веса тела в первый месяц лечения, а затем их вес возвращался к исходному и приближался к весу тела интактных животных контрольной группы. Внешний вид этих животных также отличался своеобразием, приближаясь скорее к виду нормальных животных. Шерсть была несколько реже, но волос такой же тонкий и мягкий, а изменения зубов были весьма незначительны. В этой группе погибли лишь 2 животных в первые дни лечения.
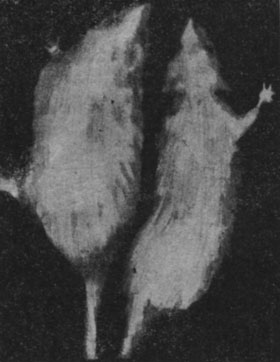
Рис. 160. - Старая крыса, получавшая эпифизигормон, по сравнению с крысой, взятой из контрольной группы
Что касается группы морских свинок, то следует отметить, что животные, подвергавшиеся тиреоидэктомии или тиреоидэктомии и кастрации, погибли самое позднее через 2 месяца после хирургического вмешательства. У этого вида животных следует подчеркнуть то обстоятельство, что вес тела оставался почти неизменным. Так, вес тела животных, подвергавшихся удалению щитовидной железы, повысился на 18% по сравнению с 26%, отмечавшимися у контрольных животных, а в группе морских свинок, подвергавшихся комбинированному удалению щитовидной и половых желез, вес тела снизился лишь на 12%. У кастрированных животных не отмечалось изменений в весе тела.
Кролики, подвергавшиеся удалению половых желез, прибавили в весе тела на 23% по сравнению с 36%, отмечавшимися у контрольных животных; кролики, подвергавшиеся удалению щитовидной железы, потеряли в весе тела 7% и, наконец, животные, у которых производилась комбинированная операция удаления как щитовидной, так и половых желез, потеряли в весе тела 10%.
В заключение можно отметить, что вес тела животных, подвергавшихся удалению щитовидной железы, снизился у всех подопытных животных, причем, наиболее значительные снижения отмечались по виду животных - у крыс, а по полу - у самцов.
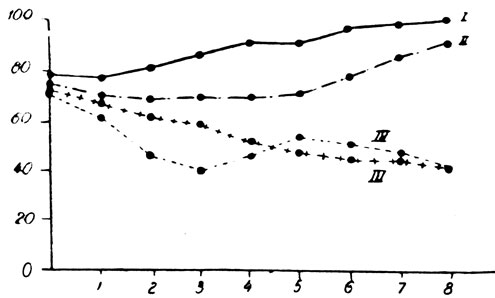
Рис. 161.- Динамика веса тела крыс-самцов, подвергнутых удалению щитовидной и половых желез, получавших эпифизгормон и витамин Е: I - Контроль; II - Животные, подвергнутые удалению щитовидной и половых желез, получавшие эпифизгормон; III - Животные, подвергнутые удалению щитовидной и половых желез, получавшие витамин Е; IV - Животные, подвергнутые удалению щитовидной и половых желез
У животных, подвергнутых удалению половых желез, отмечались различные поведенческие реакции, в зависимости от вида животного, в то время как вес тела не подвергался никакому изменению у морских свинок и слегка повышался у кроликов. У подопытных крыс разных возрастных периодов отмечались изменения в зависимости как от возраста, так и от пола животных. Так, у неполовозрелых крыс наблюдалось явное понижение веса тела, а у половозрелых и взрослых животных у самцов наблюдалась тенденция к повышению веса тела, а у самок - к снижению веса тела.
При комбинировании вышеуказанных двух железистых гипофункций отмечалась приостановка увеличения или даже снижения веса тела, причем наиболее значительные снижения наблюдались у крыс-самцов. У животных, подвергнутых двойной операции, наблюдались старческий вид пор, кожи, зубов, а также старческое поведение, что отмечалось в особенности у молодых крыс-самцов. Длительность жизни значительно укорачивалась у молодых животных всех видов, причем, максимум выживания не превышал 2-5 месяцев.
Вышеприведенные результаты указывают в некоторой степени на то, что удаление половых желез усиливает в значительной степени эффекты тиреоидэктомии, с последующим постарением животных и с укорачиванием длительности жизни.
В данном случае следует подумать также о возможном торможении регенерации тиреоидной ткани из частей ткани, оставшихся после экстирпации или из добавочной тиреоидной ткани. Следует предпринять исследования в данном направлении, тем более что отсутствие регенеративных явлений может рассматриваться как один из признаков процесса старения.
Применение некоторых препаратов, отличающихся от средств, необходимых для корригирования состояния, вызванного тиреоидэктомией и гонадэктомией, оказало благоприятное влияние на повышение веса тела, на уменьшение смертности, а также на общий вид животных: витамин Е оказал благоприятное влияние лишь на первом месяце лечения, эпифиз-гормон оказал более отчетливый эффект, продолжавшийся в течение всех 3 месяцев применения эпифизотерапии. Благоприятный эффект некоторых анаболических средств, отличающихся от медикаментозных препаратов, применяемых с целью замещения дефектов, вызванных двойной хирургической операцией, представляет серьезный интерес в отношении терапевтики старости. Это указывает, по-видимому, на то, что мы можем в известной степени воздействовать посредством различных механизмов на нарушение обмена веществ в этом возрастном периоде и таким образом достичь известных улучшений при разнообразных нарушениях старческого периода. Это вполне совпадает с тем, что как при экспериментальном лечении старости, так и при терапевтических воздействиях, применяемых в патологии человека, существует целый ряд терапевтических мер, оказывающих влияние на метаболизм в том же смысле и с теми же благоприятными эффектами.
Недавно мы предприняли ряд новых исследований, относящихся к проблеме тирео-гонадной недостаточности. Методика в общих чертах оказалась той же, что и при предыдущих опытах. Как своего рода модификацию опытов, мы ввели добавочные группы старых, оперированных животных и группы нормальных и кастрированных животных, которым вводился метилтиоурацил. Таким образом, без того, чтобы доходить до полного удаления щитовидной железы, мы старались получить определенную степень тиреоидной недостаточности, которая позволила бы наиболее длительное выживание животного. Результаты, по-видимому, подтверждают предыдущие и, кроме того, указывают на значительную смертность старых оперированных животных.
В другом ряде опытов мы исходили из идеи, что нуклеопротеиды играют очень важную роль в биологических явлениях и что эти вещества находятся в значительном количестве у детей и молодых лиц. Вилочковая железа - специфическая детскому периоду - подвергается постепенно после пубертатного периода регрессивным явлениям, несмотря на то что эту железу можно обнаружить гораздо позднее все же в довольно больших размерах (как у человека, так и у других животных); селезенка также довольно часто уменьшается в объеме у стариков, то же можно сказать и о лимфатических узлах и лимфо-эпителиальных образованиях внутренних органов. Отмечается также снижение количества лимфоцитов в крови.
Принимая во внимание все вышесказанное, мы постарались снизить количество нуклеопротеидов посредством удаления у одного и того же животного вилочковой железы и селезенки (вместо оперативного удаления можно прибегнуть к разрушению указанных органов облучением).
Опыты были поставлены на 120 животных обоего пола, а именно на неполовозрелых, препубертатного периода и взрослых крысах, а также препубертатного периода кроликах, у которых вызывалась недостаточность вилочковой железы, селезенки или обоих органов.
С целью произвести недостаточность вилочковой железы у неполовозрелых крыс мы прибегли к хирургической технике, а именно, у животных в возрасте 12 дней удалялась указанная железа, а в качестве контрольных животных мы пользовались крысятами от той же крысы-самки, с тем же весом и с рассеченной грудиной. У крыс препубертатного периода, а также у половозрелых животных вилочковая железа разрушалась при помощи рентгеновых лучей (4 сеанса по 550 г), причем сохранялись еще незначительные остатки железистой ткани. У кроликов применялась хирургическая техника.
Что касается динамики веса и роста тела, то не отмечалось никаких изменений по сравнению с контрольной группой. Не наблюдалось также никакого влияния на смертность животных. Изменения веса различных органов зависели от возраста, пола и вида животных.
При удалении вилочковой железы у крыс-самок в возрасте 12 дней наблюдается незначительное повышение веса гипофиза, надпочечников и щитовидной железы, в то время как вес половых желез слегка снижается. У самцов не было отмечено серьезных изменений.
Гистологическое исследование показало легкую гипофизарную базофилию, а также незначительное тиреопаратиреоидное и кортикосупраренальное стимулирование, более выраженное у самок. На уровне половых желез отмечается стимулирование процессов созревания. Что касается периферической крови и костного мозга, то нам не удалось отметить серьезных изменений.
У крыс в препубертатном периоде наблюдалось незначительное торможение эндокринной системы со снижением веса нижнего мозгового придатка, щитовидной железы, надпочечников и половых желез, в то время как у половозрелых животных изменения носили тот же характер, что и у неполовозрелых.
Различная реакция крыс препубертатного периода выдвигает вопрос существования переходного (критического) периода, при котором наличие вилочковой железы абсолютно необходимо, хотя нельзя исключить влияния облучения, так как животные этого возраста гораздо более чувствительны, чем взрослые, к рентгеновым лучам.
У кроликов препубертатного периода отмечалось у самок незначительное супраренально-гонадное стимулирование, в то время как у самцов наблюдалось умеренное снижение активности тех же желез, а также лимфоидная реакция селезенки. Таким образом, можно утверждать, что изменения, вызванные недостаточностью вилочковой железы, довольно незначительны и зависят от возраста и пола, причем более выражены у самок.
Недостаточность селезенки или комбинированная недостаточность как селезенки, так и вилочковой железы, изучались на неполовозрелых крысах, на крысах, находящихся в препубертатном периоде, а также на взрослых крысах. Полученные результаты встретили известные затруднения при интерпретации, так как исследование периферической крови показало у большинства животных наличие выраженной анемии с пониженным цветным показателем и с симптомами интенсивной регенерации, а миелограмма указала на гиперплазию эритробластического ростка. Точно так же отмечалось наличие эндоглобулярных паразитов типа Barthonella muris; данное паразитарное заболевание выявляется у этого вида животных при удалении селезенки. По этой причине вышеуказанные опыты были вновь проведены на других видах животных, у которых удаление селезенки не представляет этого недостатка. С вышеуказанными серьезными оговорками мы приведем все же вкратце полученные на крысах результаты.
После удаления селезенки отмечалось отставание в росте, более выраженное у самцов и у неполовозрелых животных. Относительный вес различных желез внутренней секреции - нижнего мозгового придатка, щитовидной железы, надпочечников, яичек - был повышенным, в то время как абсолютный вес находился ниже значений, отмечавшихся в контрольной группе. Гистологическое исследование указывало на умеренную гипофизарную базофилию, незначительное стимулирование щитовидной железы, а также клубочкового и пучкового слоев коркового вещества надпочечников. В отношении половых желез отмечалось торможение процесса созревания.
Наиболее значительные изменения были вызваны недостаточностью вилочковой железы и селезенки, которые изучались на неполовозрелых крысах, на находившихся в препубертатном периоде и на взрослых животных. Изменение веса отмечалось главным образом в зависимости от возраста животных. У неполовозрелых животных наблюдалось значительное снижение веса тела. Точно так же довольно выраженное снижение наблюдалось у животных в препубертатном периоде. У половозрелых животных изменения веса тела отмечались лишь после первого месяца, а затем вес тела возвращался к исходному значению.
В отличие от неполовозрелых животных, забитых через 2 месяца после удаления селезенки, половозрелые группы находились под наблюдением в течение 3-4 месяцев с целью проследить длительность выживания животных. Из группы животных препубертатного периода погибли в этот срок 13 из 16. Из половозрелых животных погибли лишь 5. Следует отметить, что у половозрелых животных были обнаружены остатки вилочковой железы, - факт, иллюстрирующий, что дозы рентгеновых лучей, предписанные для выключения функций вилочковой железы, оказались недостаточными у этого контингента животных. Следует подчеркнуть данные наблюдения о смертности, так как наши предыдущие опыты показали, что как при разрушении вилочковой железы посредством рентгеновых лучей, так и при оперативном удалении селезенки указанные вмешательства, проведенные в отдельности, не сопровождаются гибелью животных.
Вес различных органов также отличался в зависимости от возраста животных. У неполовозрелых животных отмечалось относительное повышение веса надпочечников, щитовидных желез и нижнего мозгового придатка. Однако абсолютные цифры были заниженными по сравнению с контрольной группой.
У животных в препубертатном периоде как абсолютный, так и процентный вес были занижены по отношению к контрольной группе, в то время как у половозрелых животных отмечалось повышение как относительного, так и абсолютного веса.
Гистологическая картина желез внутренней секреции у крыс с гипофункцией селезенки и вилочковой железы различалась также в зависимости от возраста. У неполовозрелых животных обнаруживалось повышение числа гипофизарных базофилов, различные картины стимулирования щитовидной железы, коркового вещества надпочечников, а также процессов полового созревания. Те же аспекты встречаются и у половозрелых животных, за исключением половых желез, где отмечаются незначительные явления инволюции. У животных препубертатного периода, с двойным хирургическим вмешательством, встречаются явления гипофизо-тирео-супрарено-гонадного торможения, причем яичник менее затронут патологическим процессом.
Однако и удаление отдельных органов могло бы привести к явлениям постарения. Это, по-видимому, происходит при удалении шишковидной железы.
Из предыдущих исследований мы могли сделать заключение, что роль шишковидной железы в организме не ограничивается лишь торможением процесса полового созревания до известного срока, характерного для различных видов, включая человеческий, а что тормозное значение этой железы распространяется на всю жизнь.
С другой стороны, исследования румынской эндокринологической школы подтвердили положение, что железа, о который идет речь, является органом с анаболическими функциями или с преимущественно анаболической деятельностью.
Удаление шишковидной железы вызывает, обычно, изменение коэффициента A/D(ассимиляция: диссимиляция) в смысле снижения ассимиляции; но это изменение приближается к явлениям, наблюдающимся при процессе старения.
Однако удаление шишковидной железы обычно вызывает более или менее выраженное поражение соседних нервных центров, что усложняет интерпретацию полученных результатов.
По вышеуказанной причине следовало выработать метод, который сводил бы к минимуму этот недостаток (Mapчела Питиш, Виктор Стэнеску и Виктор Ионеску).
На основании вышеуказанных данных мы приступили к удалению шишковидной железы у 30 крыс-самцов и самок в препубертатном периоде; мы пользовались для контроля несколькими интактными крысами или со снятой черепной покрышкой, но с сохраненной шишковидной железой (снятие покрышки черепной коробки включается в оперативный метод удаления шишковидной железы).
Указанные животные забивались спустя 2 месяца, причем исследовался главным образом эффект удаления шишковидной железы на развертывание начального периода жизни.
Другая группа, состоящая из 7 животных, подвергалась наблюдению в течение 8 месяцев, причем изучался эффект удаления железы на выживание и развитие явлений процесса старения.
В течение двух месяцев самцы с удаленной шишковидной железой прибавили в весе лишь на 25 г %, а самки на 63 г %, в то время как неоперированные контрольные животные прибавили в весе на 127 г % и соответственно на 185 г %.
Животные с удаленной черепной покрышкой, но с шишковидной железой, оставленной на месте, прибавили в весе на 68 г % и соответственно на 30 г %.
Изменение в весе эндокринных желез затрагивало главным образом половые железы, надпочечники и вилочковую железу, в то время как щитовидная железа подвергалась лишь незначительному влиянию (рис. 162).
Вес яичек увеличился в значительной степени по сравнению с контрольной группой (832 мг %; 620 мг %). У животных с простым удалением черепной покрышки вес яичек повысился лишь до 270 мг%.
Точно так же значительно повысился вес надпочечников, выразившийся в 35,7 мг%, после удаления шишковидной железы, по сравнению с 13,7 мг %, отмечавшихся у интактных животных контрольной группы, и 15 мг % - у животных с простым удалением черепной покрышки.
Вес вилочковой желзеы, наоборот, снизился у животных с удаленной шишковидной железой, равняясь 185 мг%, по сравнению с 261 мг % у интактных животных и 286 мг % у контрольной группы с простым удалением черепной покрышки.
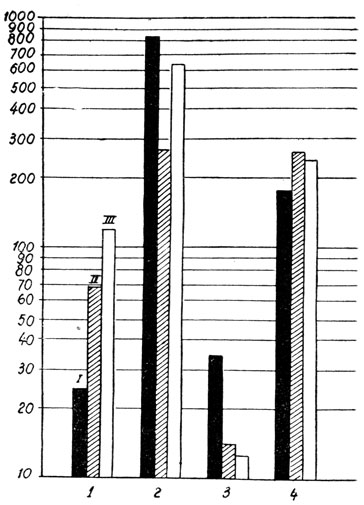
Рис. 162.- Динамика веса тела и некоторых органов крыс-самцов, подвергнутых удалению шишковидной железы по сравнению с контрольными животными: 1 - вес тела; 2 - вес яичек; 3 - вес надпочечников; 4-вес вилочковой железы. I - удаление шишковидной железы; II - оперированная контрольная группа; III - интактные животные
Точно так же снизился вес яичников до 29,3 мг % у животных с удаленной шишковидной железой, по сравнению с 44 мг % у животных контрольной группы с простым удалением черепной покрышки и 39,3 мг % у интактных животных.
Вес надпочечников самок с удаленной шишковидной железой равнялся 22,7 мг % по сравнению с 21,2 мг % у контрольных животных с удаленной черепной покрышкой и 16,5 мг % у интактных животных. Следовательно, влияние оказалось значительно менее выраженным, чем у самцов.
У самок с удаленной шишковидной железой наблюдалось повышение веса вилочковой железы, в противоположность тому, что отмечалось у самцов. Вес этого органа равнялся 400 мг % по сравнению с 220 мг % у интактных самок и 195 мг % у животных с удаленной черепной покрышкой.
С точки зрения гистологических изменений гипофиза (рис. 163 и 164) эпифизэктомированных животных отмечалось снижение количества хромофобных клеток до 52%, эозинофилов - до 18% по сравнению с 53,2% и соответственно 35,8%, отмечавшихся у контрольных животных. Базофильные клетки, наоборот, увеличились до 30% от исходных цифр 1% и соответственно 7%, наблюдавшихся у контрольных интактных и с удаленной черепной покрышкой животных. В гипофизарных эозинофильных клетках наблюдались явления гидропического набухания ядер у животных, подвергнутых удалению шишковидной железы.
В щитовидной железе (рис. 165 и 166) тех же животных отмечалось наличие отека, который, инфильтрируя соединительную ткань, сдавливал фолликулы, образовывая таким образом широкие, межфолликулярные тяжи. В менее отечных зонах тиреоидные клетки носили вакуолизированный характер протоплазмы и в большинстве случаев в них отмечались пикнотические ядра. У самок гистологические изменения были менее выражены.
Околощитовидные железы животных с удаленной шишковидной железой содержат крупные вакуолизированные клетки.
В надпочечниках наблюдались картины более выраженного стимулирования у самцов, у которых встречались значительные нагрузки клеток жировыми веществами, интенсивные расширения кровеносных сосудов, а также кровоизлияния в внешнем и среднем слоях коркового вещества надпочечника.
В яичках (рис. 167 и 168) животных, подвергнутых удалению шишковидной железы, отмечался весьма активный сперматогенез с крупными семевыводящими канальцами, диаметр которых колебался от 165 до 225 µ по сравнению с 150-180 µ у интактных животных контрольной группы и 120-150 µ у животных с удаленной черепной покрышкой. У животных, подвергнутых удалению шишковидной железы, в 88% семевыводящих канальцев наблюдались сперматозоиды, у животных контрольной группы в 4%, а у животных контрольной группы с удаленной черепной покрышкой в 0% случаев. У эпифизэктомированных животных отмечалась также значительная активность межуточной ткани. Что касается яичников, то наблюдавшиеся изменения носили весьма незначительный характер.
Изучение эпифизарных хрящей (рис. 169 и 170) бедренной и большеберцовой кости у 19 крыс, подвергнутых удалению шишковидной железы, и у 10 животных контрольной группы с удаленной черепной покрышкой показало, что у первых отмечается снижение процента энхондрального окостенения, с уменьшением количества хондроцитов, с наличием значительного количества аморфного вещества и, наконец, со сниженным участием гиалинового хряща в процессах регенерации.
Рис. 163.- Гипофиз крысы с удаленной шишковидной железой (базофилия и ядерное перерождение эозинофильных клеток)
Рис. 164. - Гипофиз крысы-самца контрольной группы
Рис. 165. - Щитовидная железа крысы-самца с удаленной шишковидной железой (отек со сдавливанием фолликулов и с вакуолизацией клеток)
Рис. 166. - Щитовидная железа крысы-самца контрольной группы
Рис. 167. - Яичко крысы-самца с удаленной шишковидной железой (активный сперматогенез по сравнению с контрольной группой)
Рис. 168. - Яичко крысы-самца контрольной группы

Рис. 169. - Эпифизарный хрящ крысы-самца с удаленной шишковидной железой
Рис. 170. - Эпифизарный хрящ крысы-самца контрольной группы
Для того чтобы уяснить, являются ли эффекты удаления шишковидной железы результатом устранения внутренней секреции указанной железы, а не раздражения или гистологически не выявляемого поражения околоэпифизарной нервной зоны, мы проследили, в какой степени парентеральное введение эпифизгормона корригирует эффекты, вызванные удалением шишковидной железы. Под нашим наблюдением находились 3 группы крыс-самцов: первая группа была подвергнута удалению черепной покрышки, вторая - удалению шишковидной железы, а крысы последней группы были подвергнуты удалению шишковидной железы с последующим парентеральным введением эпифизгормона (5 мг каждые 2 дня, в общей сложности 20 инъекций). Указанные животные забивались через 2 месяца после операции.
Мы установили, что, как и в предыдущих опытах, удаление шишковидной железы значительно изменяет вес тела, вес надпочечников и яичек, ведение эпифизгормона корригирует в значительной степени все вышеуказанные сдвиги. Что касается гистологических изменений, отмечавшихся у эпифизэктомированных животных, то последние напоминают изменения, наблюдавшиеся в предыдущей группе, и явно корригируются посредством парентерального введения эпифизарного экстракта.
В группе подвергнутых удалению шишковидной железы животных, находившихся под наблюдением в течение 8 месяцев, отмечалась более повышенная смертность оперированных животных (70%) по сравнению с контрольной группой (40%), причем у первых отмечалось наличие более редкого и более жесткого волоса. Все же, так как количество животных было незначительным, а смертность довольно высокой и в контрольной группе, мы вновь провели эти опыты на большем количестве животных. Кроме всех прочих групп, в данном случае мы добавили также группу старых эпифизэктомированных животных.
В этом отношении следует отметить опыты, проведенные нами в 1944 г. совместно с Ш. М. Милку и М. Питиш, показавшие, что парентеральное введение эпифизгормона производит благоприятный эффект на старых крыс-самцов, удлиняя их жизнь, увеличивая их вес, а также способствуя снижению старческих явлений кожи и волос. Те же результаты отмечались у старых кастрированных крыс-самок, подвергнутых лечению эпифизгормоном.
Таким образом, удаление шишковидной железы, по-видимому, укорачивает длительность инфантильного периода у крыс и приводит к более раннему созреванию, проявляющемуся в замедленном росте, половом созревании, увеличении числа гипофизарных базофильных клеток с одновременным снижением числа эозинофилов, стимулировании деятельности надпочечников и околощитовидных желез и инволюции вилочковой железы. Вышеуказанные изменения напоминают спонтанные сдвиги, появляющиеся при наступлении пубертатного периода. Изменения щитовидной железы, вызванные удалением шишковидной железы, являются в этом отношении исключением; они приводят к никогда не встречающейся у интактных животных картине, совершенно независимой от процессов, вызванных изменениями тиреотропного гормона.
Вышеуказанный фактор, сопоставленный с результатами исследований, произведенных посредством введения эпифизгормона старым животным и посредством более продолжительного наблюдения эпифизэктомированных животных, придает, по-видимому, шишковидной железе более общую роль замедлителя жизненного цикла.
Следует отметить, что эффекты удаления шишковидной железы различались в зависимости от пола, причем были ярко выражены у самцов и гораздо менее выражены у самок. Это подсказывает мысль о различном механизме осуществления половой зрелости у самцов и самок.
Для того чтобы проверить, зависит ли эффект удаления шишковидной железы на развитие жизненного цикла от наличия половых желез, мы подвергли исследованию эпифизэктомированных и кастрированных животных.
Эти опыты преследовали также и другую цель. В действительности, удаление шишковидной железы ускоряло процесс старения у крыс. С другой стороны, недостаточность или отсутствие внутренней секреции половых желез, по-видимому, приводит к тому же результату. В этих условиях нам показалось интересным исследовать, усиливает ли одновременное удаление половых желез и шишковидной железы результаты, вызывая более выраженные явления процесса старения.
Из проведенных нами опытов вытекают следующие данные:
Кастрированные, а также кастрированные и лишенные черепной покрышки крысы-самцы, весьма незначительно прибавили в весе (12 г % и 5,8 г %), в то время как животные контрольной группы с удаленной черепной покрышкой и с сохраненными половыми органами прибавили в весе 59,4 г %, животные, подвергнутые удалению шишковидной железы, прибавили в весе 41,7 г %, а кастрированные и эпифизэктомированные животные прибавили в весе 35 г%.
У самок различия между разными группами значительно менее ярко выражены. Так, в контрольной группе с удаленной черепной покрышкой отмечалось повышение веса тела на 53,9 г %, у эпифизэктомированных животных прибавление в весе тела равнялось 56,7 г %, а у животных, подвергнутых двойной хирургической операции, составляло 45,9 г %, в то время как у кастрированных и лишенных черепной покрышки животных прибавление в весе равнялось 31,3 г %, а у только кастрированных составляло 27,5 г %. Следовательно, и в данных опытах при кастрации наблюдалось самое значительное приостановление развития веса тела.
Что касается веса отдельных органов, то следует отметить, что гипотрофия гипофиза, вызванная кастрацией, оказывает менее выраженное влияние на эпифизэктомированных животных: 11,8 мг % у самцов и 8,4 мг % у самок по сравнению с 13 мг% и соответственно 10 мг % у кастрированных самцов и самок с сохраненной шишковидной железой.
В отношении надпочечников наблюдалось различное поведение в зависимости от пола: 19,6 мг % у кастрированных самцов; 21,5 мг % у самцов с удаленной шишковидной железой и 20,4 мг % при применении комбинированной операции. У самок 17,2 мг % после кастрации, 21,5 мг % после удаления шишковидной железы и 18,1 мг % при применении комбинированной хирургической операции. У животных контрольной группы с удаленной черепной покрышкой - 19,5 мг %.
Семенные пузырьки весили 31 мг% у кастрированных самцов по сравнению с 53,4 мг %, наблюдавшимися у кастрированных и эпифизэктомированных животных. Удаление шишковидной железы вызывает, следовательно, повышение веса семенных пузырьков даже при отсутствии яичек.
Гистологические изменения после удаления шишковидной и половых желез различаются в зависимости от пола: в то время как первые преобладают у самцов, сдвиги, вызванные кастрацией, более выражены у самок.
Из этих исследований вытекает, что влияние шишковидной железы не выполняется посредством половых желез, так как изменения, вызванные удалением шишковидной железы (уменьшение количества и дегенеративные изменения гипофизарных эозинофилов, отек щитовидной железы и т. д.), встречаются также у кастрированных и эпифизэктомированных животных, а различное действие в зависимости от пола наблюдается и у кастрированных животных. Следует все же принять во внимание возможное влияние удаления шишковидной железы на андрогенную ткань надпочечников.
Что касается смертности и явлений процесса старения, то нам не удалось отметить ничего особенного, во всяком случае в тот короткий промежуток времени (2 месяца), в который животные, подвергнутые комбинированной операции, находились под наблюдением.
В продолжение исследования роли двужелезистой недостаточности в явлениях старости и в особенности в наступлении пубертатного периода ставились опыты на неполовозрелых крысах-самцах. Через 10 дней после удаления эпифиза животные получали в течение 50 дней метилтиоурацил по 10 мг в сутки. Контролем служили животные, получавшие и неполучавшие метилтиоурацил.
Повышение веса тела крыс с удаленной шишковидной железой и с последующим введением метилтиоурацила равнялось 69%, а в контрольной группе с введением метилтиоурацила оно равнялось 93,8%, в то время как в контрольной группе без применения указанного средства вес тела животных повышался на 126,7%. Вес щитовидной железы значительно отличался у обработанных крыс (эпифизэктомированных или интактных) и необработанных; то же самое можно сказать относительно веса надпочечников эпифизэктомированных и обработанных крыс по сравнению с весом надпочечников крыс контрольной группы.
Гистологическое исследование и весовые данные показали, что при введении метилтиоурацила сдвиги, вызванные удалением шишковидной железы в гипофизе, надпочечниках и половых железах более стертые; это указывает на то, что появление ранней половой зрелости при удалении шишковидной железы требует присутствия определенного количества тиреоидного гормона, что вполне совпадает с дифференцирующей ролью, приписываемой щитовидной железе.
Мы исследовали также двужелезистую эпифизо-супраренальную недостаточность и пришли к заключению, что удаление шишковидной железы не изменяет длительности жизненного цикла супрареналэктомированных животных. Разумеется, в этом отношении нужно было бы исследовать эпифизэктомированных животных с частичной недостаточностью надпочечников.
В другой главе мы указали на роль, которую играет кора головного мозга в процессе старения, подтвержденную целым рядом клинических и экспериментальных данных. Во время произведенных нами опытов по изучению проблемы невроэндокринных соотношений* отмечался целый ряд трофических расстройств, напоминающих нарушения, наблюдаемые при старении у кошек с двусторонним поражением глазничной области. У двух кошек с вышеуказанным поражением, проанализированном 6 месяцев после хирургического вмешательства, мы отметили прогрессирующую кахексию с трофическими расстройствами кожи и шерсти, с редким волосом, с островками оголенной кожи, с шелушением, с пятнистыми витилигообразными поражениями, с обезвоживанием и потерей эластичности покровов. У одного из животных отмечалась полная тестикулярная инволюция, с замещением тестикулярной ткани склеротической (рис. 171 и 172). У другого животного с тем же поражением через месяц после хирургического вмешательства мы отметили начало перерождения яичек с склеиванием сперматидов и с отсутствием сперматозоидов. Принимая во внимание незначительное количество случаев, нельзя сделать обобщающее заключение; обсуждение проведенных нами наблюдений можно будет сделать лишь после окончания текущих опытов (рис. 173).
* (Исследования были проведены совместно с М. и К. Бэлэчану.)
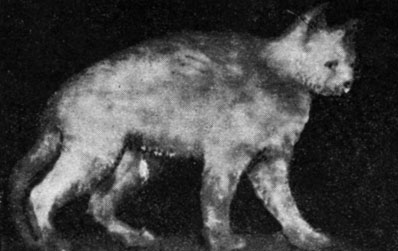
Рис. 171. - Кот с двусторонним поражением глазничной области, с трофическими расстройствами, напоминающими старческие нарушения у кошек
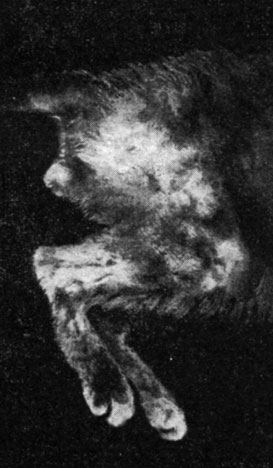
Рис. 172. - Внешний вид кожи и шерсти того же животного, что и на рис. 171

Рис. 173. - Яичко кота с двусторонним поражением глазничной области
У неполовозрелых децеребрированных крыс развитие жизненного цикла видоизменяется так, как было указано в наших опытах*. Яички животных с удаленной корой весили 528,7 мг % по сравнению с 363 мг % у контрольных животных с удаленной черепной покрышкой. Что касается яичников, то нам не удалось отметить значительных изменений в их все; в отличие от эпифизэктомированных животных с декортикацией, животные выросли значительно больше, чем животные контрольной группы с удаленной покрышкой черепа, причем у последних не отмечалось достоверных изменений надпочечников. Гистологическое исследование показало прилив крови и инфильтрацию всех эндокринных желез с ранним созреванием семенных пузырьков, но не отмечалось изменений, подобных вызванным удалением шишковидной железы, в гипофизе, в щитовидной железе и в надпочечниках.
* (Исследования были проведены совместно с М. Питиш, В. Стэнеску и В. Ионеску.)
Если в до сих пор приведенных опытах мы исследовали экспериментальное наступление процесса старения, то в другом ряде опытов мы занимались исследованием лечения старости у белых крыс и подчеркнули, что в общих чертах процесс старения обратим, и таким образом длительность жизни может быть увеличена. Указанные исследования* имели целью изучение терапевтического эффекта, полученного в результате введения экстракта вилочковой железы, экстракта легких, пантотеновой кислоты и антиретикулярной цитотоксической сыворотки Богомольца старых крыс.
* (Проведенные совместно с М. Питиш.)
Эти опыты являются продолжением прежних исследований, в которых мы изучали эффекты введения фолликулина, тестостерона, вытяжки щитовидной железы, инсулина, эпифизарного экстракта, жирорастворимого овариального экстракта и т. д. на явления постарения и длительности жизненного цикла у крыс.
Лечению подвергались специально отобранные крысы старческой группы, с общей слабостью, с почти полным выпадением шерсти, с утолщенной и грубой кожей, причем у некоторых из животных наблюдались также изъязвления покровов. Что касается психического поведения, то у указанных животных отмечалось апатическое состояние и безразличное отношение к каким бы то ни было раздражителям. Мы исследовали эффект, произведенный на указанные поражения, а также на длительность жизненного цикла, весовые изменения и на рост (принимая во внимание наличие эпифизарных хрящей у этого вида животных).

Рис. 174. - Старая крыса, подвергнутая лечению экстрактом вилочковой железы
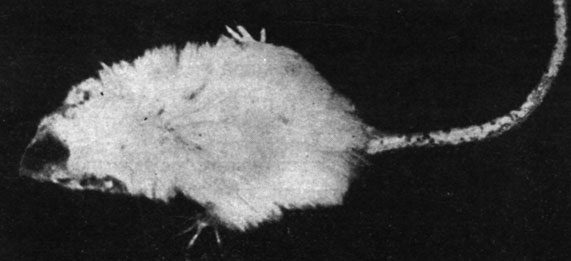
Рис. 175. - Старая крыса контрольной группы
Лечение экстрактом вилочковой железы оказалось с самого начала весьма обнадеживающим, поскольку тимус является железой детского периода, играющей значительную роль в анаболическом и гидратирующем метаболизме, будучи весьма богатой нуклеопротеидами (рис. 174 и 175).
Мы вводили водорастворимый экстракт, полученный посредством щелочного лизиса вилочковой железы, в количествах 0,5 мл, каждые два дня, в течение 5 месяцев.
После первого месяца лечения погибли 2 крысы контрольной группы и 1 крыса, которой делались инъекции. Вес тела подвергнутых лечению животных повысился в среднем на 18,6%, в то время как вес контрольных животных снизился в тот же промежуток времени на 5,2%.
Различия между инъецированными и контрольными животными еще более усилились на второй месяц исследований. Так, в этот период умерла одна контрольная крыса, а вес тела животных, подвергнутых лечению, повысился в среднем на 43,2% по сравнению и исходным, в то время как вес тела контрольных животных повысился лишь на 10,3%.
Сопоставляя прибавку в весе тела инъецированных животных с прибавкой в весе тела животных контрольной группы, можно отметить, что первые повысились в весе на 35% по сравнению с последними.
В течение третьего месяца погибли еще 2 крысы контрольной группы, в то время как в группе подвергнутых лечению животных не отмечалось ни одного смертельного исхода. Средняя веса тела крыс, подвергнутых лечению, повысилась на 48,1% по отношению к исходному весу, а у контрольных животных лишь на 8%, следовательно, отмечалось относительное снижение в отношении предыдущего месяца. Инъецированные животные прибавили за эти 3 месяца в весе на 60% по сравнению с контрольными крысами.
В течение четвертого месяца погибли последние 3 контрольные крысы и одна из подвергнутых лечению. В этот месяц вес тела животных претерпел незначительное снижение, причем средняя прибавки веса тела равнялась 38,2% по сравнению с исходным значением.
В течение пятого месяца лечения погибло еще одно из подвергнутых лечению животных.
Средняя прибавки в весе тела равнялась лишь 30,6%, следовательно, отмечалась тенденция к снижению, хотя вес животных был более завышенным по сравнению с исходной величиной.
Кроме прибавки в весе тела, инъецированные животные выросли в длину, что вполне объяснимо даже у старых крыс, так как у этого вида животных не отмечается исчезновения эпифизарных хрящей в продолжение всей жизни. Следует подчеркнуть, что при соответствующих анатомических условиях рост в длину может вновь появиться даже у очень старых животных. У животных, подвергнутых лечению, отмечался также рост шерсти, изменение внешнего вида покровов, ставших более эластичными, исчезновение изъязвлений, а также изменение поведения в смысле исключительной живости животных. Изменение внешнего вида носило прогредиентный характер в первые три месяца и сохранилось в том же состоянии в последние 2 месяца.
Таким образом, можно утверждать, что экстракт вилочковой железы вызвал у крыс явную регрессию многочисленных явлений старения и продлил их жизненный цикл.
Еще в 1948 г. мы изложили причины, заставившие нас утверждать, что дыхательная система включается в общую биологическую деятельность не только посредством функции введения кислорода и выведения углекислоты, но и посредством гуморального механизма в обменных явлениях, в частности, в феноменах, находящихся в прямом соотношении с функцией тканевого дыхания. В этой связи было бы очень интересно исследовать, может ли дыхательная система посредством вышеуказанного гуморального механизма оказать влияние на длительность жизни; большинство исследователей старается связать длительность жизненного цикла с окислительными явлениями и даже с объемом легких.
Мы изучали эффект, произведенный легочным экстрактом, приготовленным из легких лошади посредством щелочного лизиса; отсепарирование бронхиол не производилось полностью, так что в дальнейшем нужно будет изучить деятельность как альвеолярной, так и бронхиальной ткани. В ряде весьма интересных работ Поликар вполне обоснованно настаивал на структурных различиях указанных тканей. В течение 4 месяцев мы вводили указанный экстракт в количестве 0,5 мл каждые два дня 12 старым крысам (10 самцам и 2 самкам), в то время как 12 животных сохранялись в качестве контрольной группы. Через несколько дней после опыта погибли 3 животных контрольной группы.
В течение первого месяца лечения погибли 2 контрольных крысы и 1 инъецированная крыса; вес тела снизился у животных, подвергнутых лечению, на 3%, а в контрольной группе на 3,5%, причем диапазон колебаний был весьма незначительным у контрольных животных (от 2 до 9%) и довольно значительным у инъецированных крыс (у 7 из последних изменения колебались от 6 до 30%), а у остальных 5 животных вес тела оставался стационарным или даже слегка повышался.
На второй месяц средняя веса тела животных, подвергнутых инъекциям, снизилась на 12% по сравнению с исходным весом, а у животных контрольной группы повысилась на 2%.
На третий месяц погибло 3 контроля и 6 животных, подвергнутых инъекциям. Так как осталось слишком незначительное количество животных (самых крепких и выносливых), нам трудно интерпретировать весовые изменения. Все же было установлено, что вес тела инъецированных животных продолжал снижаться (16%), а вес тела в контрольной группе оставался стационарным или даже слегка увеличивался.
На четвертый месяц погибли и последние 2 инъецированные крысы, вес тела которых продолжал прогрессивно снижаться. Нам не удалось отметить улучшений во внешнем виде или поведении животных; рост последних оставался также не измененным.
Неблагоприятный эффект легочного экстракта у старых крыс можно было, по-видимому, объяснить его катаболическим и обезвоживающим действием, подтвердившимся и при других экспериментальных работах и клинических наблюдениях, причем в этом отношении известную роль играет, вероятно, стимуляция щитовидной железы. Возможно, мы добились бы других результатов при другой дозировке.
Учитывая сходство многочисленных феноменов гипо- или авитаминозов и расстройств, появляющихся в старческом периоде, а также важную роль, которую играют указанные вещества в ферментных системах организма, мы посчитали необходимым исследовать терапевтический эффект не только гормонов, но и других эргонов, в частности витаминов. Значительная роль пантотеновой кислоты в предупреждении и лечении ахромотрихоза, а также значение, приписываемое этому витамину в тканевом питании и в жировом и углеводном обменах, заставили нас исследовать действие этой кислоты на старых животных. Рассматривая поседение волос и некоторые феномены гипофункции надпочечников как симптомы процесса старения, мы задались целью добиться омоложения животных посредством указанного лечения.
Лечение проводилось в течение 50 дней в группе, состоящей из 16 старых крыс, самцов и самок, посредством парентерального введения 1 мл ежедневно (0,025 г пантотеновой кислоты).
Что касается клинической стороны проблемы, то можно отметить, что у подвергнутых лечению животных наблюдались известные признаки омоложения: рост шерсти, более гладкая и более эластичная кожа, по сравнению с контрольной группой.
Вес тела животных изменился довольно незначительно, - прибавка в весе по сравнению с контрольной группой равнялась примерно 5-10%. Следует, однако, отметить, что животные реагировали на указанное лечение различно в зависимости от пола: более значительные изменения наблюдались у самок.
Отмечалась выраженная прибавка в весе эндокринных желез - гипофиза, шишковидной железы, щитовидной железы и надпочечников, и менее выраженное повышение веса различных органов: печени, сердца, головного мозга, причем наиболее значительное повышение веса отмечалось у самок.
Указанному повышению веса соответствуют гистологические изменения: стимуляция хромофобных клеток гипофиза с пропорциональным снижением клеток хромофильного типа, стимуляция клеток шишковидной железы, доходившая до появления зон разрастания, стимуляция щитовидной железы с наличием клеток экскреторного типа. Что касается яичек, то отмечалась стимуляция сперматогенеза с наличием многочисленных кариокинезов, в то время как межуточная ткань оставалась неизменной. В яичниках наблюдалась активация фолликулов, в особенности их раннее созревание, и активация межуточных клеток с появлением зон разрастания. В вилочковой железе не отмечалось различий по отношению к контрольным самкам, а что касается инъецированных самцов, то в данном случае уменьшение объема железы, как и уменьшение количества лимфоцитарных элементов можно было бы интерпретировать как влияние активной тестикулярной ткани. Что касается различных внутренних органов, то нам не удалось отметить никаких достоверных сдвигов по сравнению с контрольной группой (рис. 176-189).
Исследования А. А. Богомольца явились исходным пунктом для испытаний другого терапевтического метода, примененного на старых крысах.
А. А. Богомолец, занимаясь изучением соединительной ткани, утверждал, что последняя не является простой тканью, заполняющей пустые пространства, а наоборот, ее клетки играют активную роль в обмене, происходящем между специфическими клетками различных тканей и капиллярами, прекапиллярами и т. д. Этот видный биолог рассматривает соединительную ткань как "основу" организма.
Исходя из этой идеи, вышеупомянутый автор приготовил сыворотку из крови животных, инъецированных экстрактами селезенки и костного мозга - органов, содержащих большое количество ретикуло-эндотелиальных соединительных клеток.
При помощи этой антиретикулярной цитотоксической сыворотки, применявшейся в небольших количествах, ему удалось достичь активации старческого организма, увеличения регенеративной способности организма, ускорения срастания переломов, повышения устойчивости по отношению к раку и т. д.
В концепции А. А. Богомольца о роли активных клеток соединительной ткани, эндотелия капилляров включительно, не учитывалось значение учения И. П. Павлова о роли коры головного мозга в функциях целостного организма.
С другой стороны, следует принять во внимание роль различных рецепторов, находящихся в глубине ткани, включая роль рецепторов активной соединительной ткани, которые посылают в кору головного мозга импульсы, появляющиеся в области указанных тканей, и, таким образом, обеспечивают единство организма.
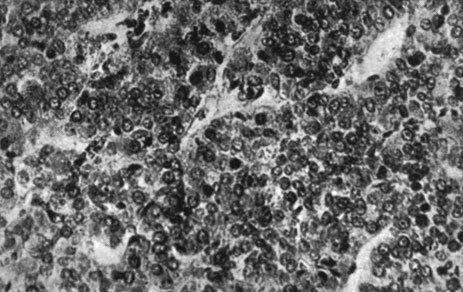
Рис. 176. - Гипофиз старой крысы, подвергнутой лечению пантотеновой кислотой (видно хромофобное разрастание)
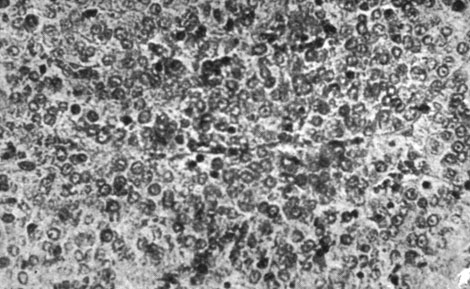
Рис. 177. - Гипофиз старой крысы контрольной группы
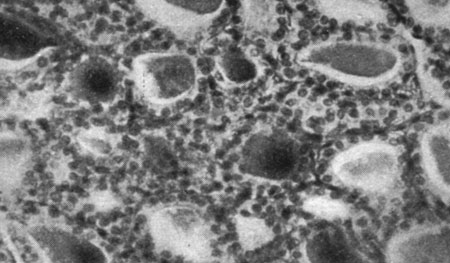
Рис. 178. - Щитовидная железа старой крысы, подвергнутой парентеральному введению пантотеновой кислоты
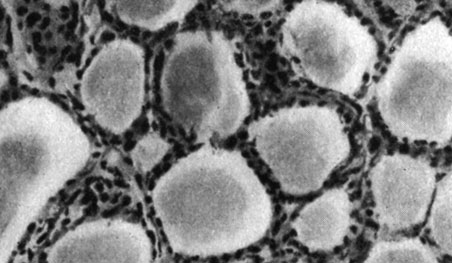
Рис. 179. - Щитовидная железа старой крысы контрольной группы
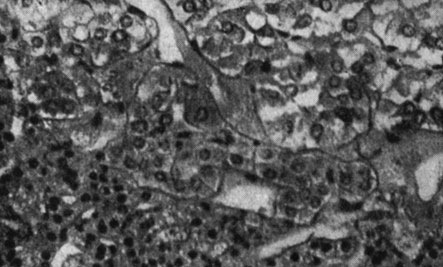
Рис. 180. - Надпочечник старой крысы, подвергнутой парентеральному введению пантотеновой кислоты
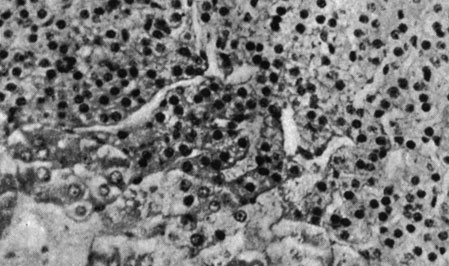
Рис. 181. - Надпочечник старой крысы контрольной группы
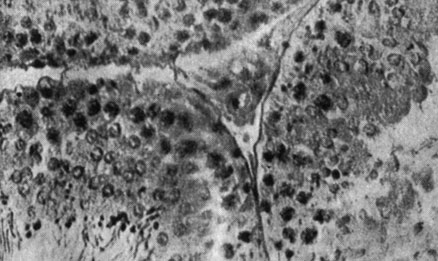
Рис. 182. - Яичко старой крысы после парентерального введения пантотеновой кислоты
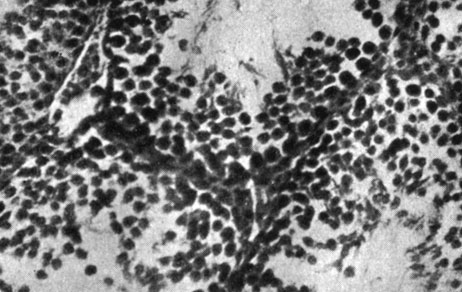
Рис. 183. - Яичко старой крысы контрольной группы
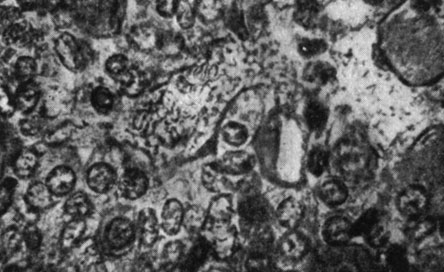
Рис. 184. - Гипофиз старой крысы, подвергнутой введению сыворотки Богомольца
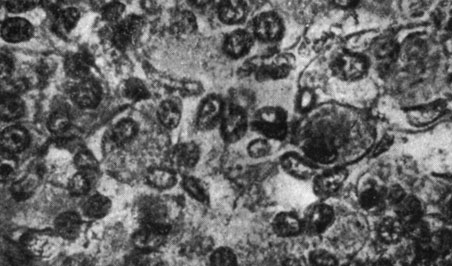
Рис. 185. - Гипофиз старой крысы контрольной группы
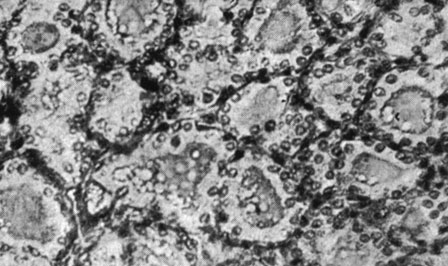
Рис. 186. - Щитовидная железа старой крысы, подвергнутой введению сыворотки Богомольца
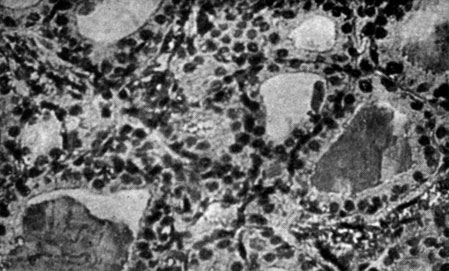
Рис. 187. - Щитовидная железа старой крысы контрольной группы
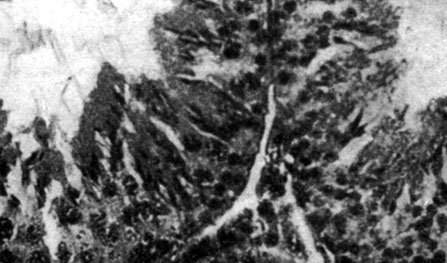
Рис. 188. - Яичко старой крысы, подвергнутой введению сыворотки Богомольца
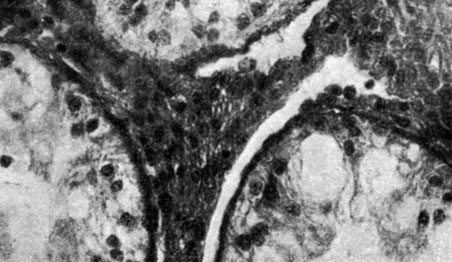
Рис. 189. - Яичко старой крысы контрольный группы
С этими оговорками мы считаем, что исследования А. А. Богомольца, в связи с его антиретикулярной цитотоксической сывороткой (приготовленной посредством парентерального введения размельченной селезенки и костного мозга), большие дозы которой производят токсическое действие, а малые оказывают стимулирующее действие, заслуживают внимания.
Мы лично предприняли ряд экспериментальных исследований, относящихся к действию этой сыворотки на старых крыс. Приводим ниже наиболее интересные результаты.
Старые крысы - самцы и самки - были разбиты на группы по 8 животных. Сыворотка А. А. Богомольца вводилась парентерально каждые 2 дня в количестве 0,2 мл, в разведении 1/10, в течение 50 дней. Во втором ряде опытов мы применяли на группе, состоящей из меньшего числа животных (4 крысы), сыворотку румынского производства, приготовленную по тому же методу в Институте им. И. К. Кантакузино.
Что касается изменений веса тела, то нам не удалось отметить значительных различий. Гистологическое исследование показало активацию всей эндокринной системы. Это выражается на уровне передней доли гипофиза в наличии главных (хромофобных) клеток и зон разрастания, а также в уменьшении количества базофильных клеток с параллельным ростом эозинофильных. Наличие клеток юного типа в межуточной и задней долях гипофиза, а также двуядерных клеток в шишковидной железе следует рассматривать в том же смысле.
Активация щитовидной железы выражается в микрофолликулярной структуре и в наличии вакуолей, а также многочисленных клеток экскреторного типа.
В области околощитовидных желез отмечается также наличие зон разрастания с преобладанием ацидофильных клеток, которые считаются активными. Надпочечник подвергается известным изменениям: в то время как у животных контрольной группы жировые вещества встречаются во всех слоях, главным образом в сетчатом, у подвергнутых лечению животных сетчатый слой образован из зернистых клеток с зонами разрастания. В мозговом веществе железы отмечается активная картина с хромаффинными клетками секреторного типа.
В области яичек отмечается также интенсивная активация. В яичниках обнаруживается тенденция к лутеинизации.
Опыты, относящиеся к лечению процесса старения, следует умножить, учитывая наличие различных видов животных, а также различных способов лечения. Кроме того, они должны длиться более или менее продолжительное время, чтобы установить, в какой степени определенное терапевтическое средство теряет свое действие после определенного промежутка времени. Затем выдвигается вопрос комбинирования различных терапевтических методов, одновременно или последовательно, что, возможно, увеличивает интенсивность или поздний эффект данного лечения. Возраст, когда начинается лечение - также один из факторов, который в состоянии повлиять на длительность жизни различных видов.
Индивидуальное развитие может быть изменено, если мы применим терапевтическое вмешательство в различные стадии развития. Мы видели, что известные терапевтические методы или вмешательства, применяемые на старых, взрослых или молодых животных, могут ускорить процесс старения, замедлить последний или даже повлиять в благоприятном смысле, другими словами, вызвать омоложение животного.
Терапевтическое вмешательство, применяемое в очень ранних стадиях - во время эмбрионального развития - дает наибольшие возможности повлиять на последующее развитие индивида, и все же данные возможности были до сих пор недостаточно изучены. Указанные методы терапии представляют собой наиболее обещающие области экспериментальной иликибиологии.
Много лет тому назад нами была выдвинута гипотеза о том, что железы внутренней секреции функционируют и в течение зародышевого развития организма.
В монографии, посвященной этим функциям, Les secretions internes, вышедшей в свет в 1909 г., мы отмечали следующее: "Вопрос ахондроплазии выдвигает проблему вмешательства желез внутренней секреции в течение зародышевой жизни, причем соответствующая эндокринная функция может быть, a priori, настолько же необходимой, как и в течение внеутробной жизни"*.
* (С. I. Parhon, M. Goldstein, Les secritiones internes, Maloine, Париж, 1909, стр. 309.)
Мы вновь вернулись к этой проблеме в 1913 г., настаивая на роли различных желез в органообразовании и гистообразовании.
Мы показали, что некоторые симптомы аплазии щитовидной железы приводят к приостановке развития в определенный период зародышевой жизни и заслуживают названия "фетализма" - термин, применявшийся нами еще в работе 1909 г.
Наиболее наглядным нам показался пример молочной железы. Она отсутствует или находится лишь в зародышевом состоянии у практически здоровых мужчин и у детей обоего пола. Она появляется у женщин и самок млекопитающих в пубертатном периоде, причем ее образование строго обусловлено внутренней секрецией яичников. Кастрация препятствует образованию молочной железы, которая нормально развивается лишь в случае, если удаление яичников компенсируется успешной подсадкой указанных желез. Это является весьма показательным примером, указывающим на роль эндокринных желез в образовании органов.
Можно было бы привести и другие примеры: яичники, например, влияют на развитие матки.
Можно предположить, что в указанные периоды развития, природа не прибегает к различным в основе механизмам, и таким образом мы имеем все основания утверждать, что во время внутриутробной или, в более широком смысле слова, в период зародышевой жизни, когда главным образом образуются органы, эндокринные железы вступают в действие в наиболее активной форме.
Что касается метода исследования проблемы, то мы уже отмечали что проведение опытов в этой области встречает значительные затруднения, но все же можно было бы провести исследования на зародышах рыб или лягушек.
Разумеется, в настоящий момент мы находимся в гораздо более благоприятных условиях в отношении методики экспериментальных исследований.
В 1913 г. мы упоминали, что другой методикой могло бы явиться изучение тератологических случаев с целью установить, существует ли взаимоотношение между отсутствием развития какого-либо органа и другими органами. В этом отношении мы напоминали о наличии сосуществования анэнцефалии с аплазией надпочечников и в связи с этим сосуществованием выдвигали гипотезу, защищенную нами в 1910 г., что аплазия вышеуказанных желез, по-видимому, обусловлена аплазией мозга. Клетки коры головного мозга во время своего развития сецернируют гормон, стимулирующий корковое вещество надпочечника, а последний сецернирует в свою очередь вещества, необходимые развитию головного мозга.
Чередование развития органов следовало бы также принять во внимание, так как ранее развитые органы создают, вероятно, условия, необходимые развитию позднее появляющихся органов.
К этому добавим, между прочим, что гормоны зародышевой пищеварительной трубки могут, по-видимому, принимать участие в образовании придаточных пищеварительных желез.
Культуры, выращенные in vitro в плазме, с прибавкой экстрактов или зародышевых тканей, могли бы тоже дать определенные результаты.
В работе, вышедшей в свет в 1923 г., мы вернулись вновь к этим проблемам, настаивая главным образом на гистогенетической роли гормонов, а также на возможности существования общей закономерности в том смысле, что между некоторыми органами или тканями, из которых одни стимулируют развитие других, возникают антагонистические соотношения после завершения их развития, причем ранее стимулирующий орган позднее становится тормозящим. Так можно было бы объяснить аменоррею женщин, страдающих тяжелой формой макромастии.
В последнее время мы вновь вернулись к проблеме роли гормонов в эмбриогенезе. Недавно мы опубликовали результаты исследований, проведенных на куриных зародышах, посредством инъекций эпифизарного экстракта в хориоаллантоисную оболочку или в амниотическую полость, причем получили активацию роста, раннее и более быстрое развитие оперения и повышенную устойчивость к различным патологическим факторам, например к авитаминозу (рис. 190 и 191).
Затем мы исследовали роль тироксина в эмбриогенезе кур. Известно, что тироксин ускоряет превращение головастика в лягушку, вызывает метаморфозу аксолотля, который в нормальных условиях является педогенетическим и, наконец, в определенных дозах оказывает влияние на рост млекопитающих и даже человека.
Зная указанное воздействие тироксина на рост и дифференцировку, мы решили, что сможем ускорить зародышевое развитие кур и, таким образом, укоротить инкубационный период; полученные результаты не соответствовали, однако, нашим ожиданиям.
Мы подвергли куриные зародыши породы леггорн и род-айланд действию раствора тироксина, вводимого в хориоаллантоис. Обработка проводилась ежедневно между 5-м и 15-м днем развития посредством инъекций на нескольких группах яиц. Общая доза, вводившаяся зародышу, колебалась в пределах от 1 до 5,5 у тироксина. Соответствующие контрольные животные получали такие же дозы физиологического раствора.
В этих опытах нам удалось получить 14 цыплят, вылупившихся спонтанно; при вылуплении у цыплят наблюдались лишь весьма незначительные изменения в весе тела по сравнению с контрольной группой. Часть цыплят погибла от интеркуррентных заболеваний вскоре после вылупления, так что под нашим наблюдением осталось лишь 5 цыплят, которых мы смогли проследить в динамике. У этих цыплят наблюдался карликовый рост, усилившийся одновременно с возрастным развитием. Рентгенограммы эпифизарных хрящей указывали на менее значительно развитые точки окостенения, чем у животных контрольной группы, и более широкие хрящевые зоны, что продолжалось довольно долго; в то время как окостенение развивалось вполне нормально в контрольной группе, у подвергнутых обработке цыплят отмечалась задержка последнего. Указанное замедление роста происходило параллельно с динамикой веса тела. Так, например, после 3 месяцев цыплята, подвергнутые обработке, весили 175-260 г, в то время как вес цыплят контрольной группы колебался в пределах 400-600 г (рис. 192-201).
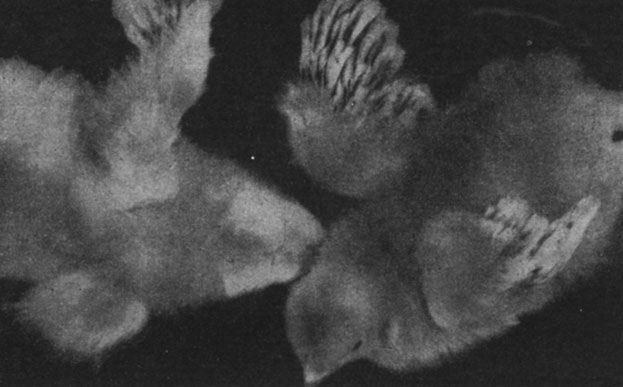
Рис. 190. - Цыплята в возрасте 18 дней. Слева контроль, справа цыпленок, подвергнутый внутризародышевой обработке шишковидной железой (видно более выраженное развитие оперения)

Рис. 191. - Цыплята в возрасте 30 дней. Слева контроль, справа цыпленок, подвергнутый лечению (видно более выраженное оперение)
Гистологическое исследование желез внутренней секреции этих цыплят не показало статистически достоверных изменений. Отмечалось, однако, гипофизо-тирео-супраренальная гипертрофия. Гелэн-Шедрина подвергла также обработке тироксином развивающихся куриных зародышей, но ей удалось получить лишь зародышей в возрасте 11-12 дней, у которых отмечались гиперемия и гипертрофия органов, атрофия селезенки и инфильтрация кожи, главным образом, в области головы и живота - явления, отмечавшиеся и нами у мертвых цыплят в инкубационном периоде; по нашему мнению, указанные явления не специфичны.

Рис. 192. - Цыплята в возрасте 12 дней. Слева контроль, справа цыпленок, подвергнутый внутризародышевой обработке тироксином

Рис. 193. - Цыплята в возрасте 40 дней. Слева цыпленок, подвергнутый внутризародышевой обработке тироксином, справа контроль
Рис. 194. - Цыплята в возрасте 70 дней. Слева контроль, справа 2 цыпленка, подвергнутые внутризародышевой обработке тироксином
Рис. 195. - Цыплята в возрасте 70 дней. Слева контроль, справа цыпленок, подвергнутый обработке (видно бедное оперение, короткие и тонкие перья)
М. С. Мицкевич, инъецируя в голубиное яйцо антитиреоидные вещества (метилтиоурацил), получил гипертрофию щитовидной железы с вакуолизацией фолликулярных клеток, задержку развития с сохранением желточного мешка и замедление вылупления.
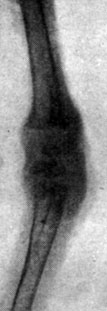
Рис. 196. - Цыпленок контрольной группы в возрасте 34 дней. Рентгенограмма эпифизарных хрящей коленного сустава
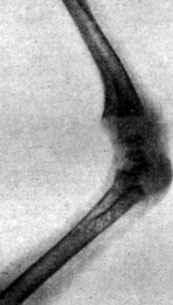
Рис. 197. - Цыпленок в возрасте 34 дней, подвергнутый внутризародышевой обработке тироксином. Рентгенограмма эпифизарных хрящей коленного сустава
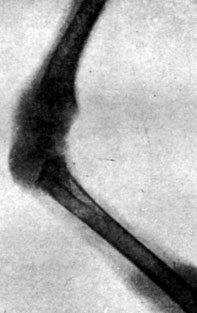
Рис. 198. - Цыпленок контрольной группы в возрасте 41 дня. Рентгенограмма эпифизарных хрящей коленного сустава
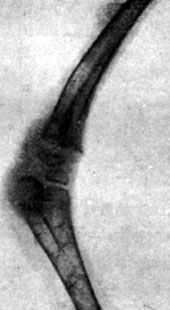
Рис. 199. - Цыпленок в возрасте 41 дня, подвергнутый внутризародышевой обработке тироксином. Рентгенограмма эпифизарных хрящей коленного сустава
Полученные нами результаты на первый взгляд кажутся аналогичными с наблюдающимися у головастика лягушки при ускорении созревания под воздействием тироксина, но с одновременной задержкой роста. В наших опытах мы, однако, не отмечали укорочения инкубационного периода, а эпифизарные хрящи, изучавшиеся в более позднем периоде, занимали еще широкую зону, в то время как точки окостенения были весьма незначительными, что исключает гипотезу блокады эпифизарных хрящей посредством введения слишком завышенных доз тироксина.
Возможно, что введенный в организм тироксин приостановил деятельность щитовидной железы зародыша и вызвал, таким образом, карликовый рост как результат гипофункции щитовидной железы. Щитовидная железа зародыша цыпленка дифференцируется морфологически спустя 48 часов, а по некоторым исследованиям она становится функционально активной через 10 дней, когда можно установить накопление йода в данном органе. Следовательно, введенный тироксин нашел соответствующий приемник и, таким образом, стала вероятной гипотеза приостановки деятельности щитовидной железы. Однако у цыплят не наблюдалось микседематозного аспекта; они были бойкими, подвижными, а что касается морфологии щитовидных желез, то нам удалось отметить лишь незначительную гипофункцию последних.
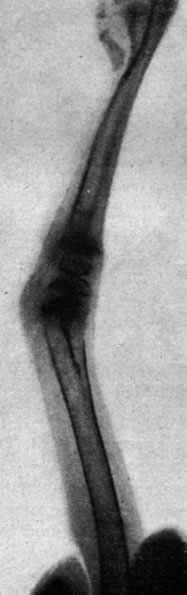
Рис. 200. - Цыпленок в возрасте 60 дней контрольной группы. Рентгенограмма коленного сустава
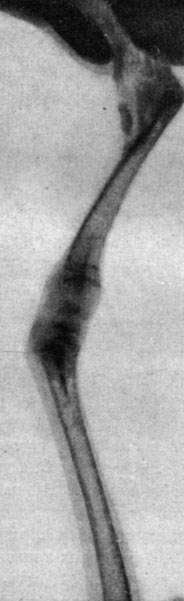
Рис. 201. - Цыпленок в возрасте 60 дней, подвергнутый внутризародышевой обработке тироксином. Рентгенограмма коленного сустава
Возможно также, что тироксин действовал как общее токсическое средство и вызывал, таким образом, карликовый рост токсического происхождения. Однако нам не удалось обнаружить поражений в других органах, несмотря на то, что надпочечники оказались увеличенными, вероятно, вследствие прямого влияния тироксина, что хорошо известно из литературы по специальности.
Полученный таким путем карликовый рост весьма напоминает нанизм, вызванный гипофизарной недостаточностью: речь идет о гармоническом нанизме, усиливающемся со временем и который можно было воспроизвести торможением нижнего мозгового придатка посредством введения тироксина. Это объяснило бы и общий тиреоидный аспект. Н. А. Шерешевский рассматривает нанизм гипофизарного происхождения как неврогипофизарную дистрофию, и действительно, это могло бы произойти при токсическом воздействии тироксина на центральную нервную систему. В клинике мы наблюдали случаи заболевания головного мозга, вызванные повышенной концентрацией тироксина в крови. Этот неврогипофизарный механизм является, по нашему мнению, наиболее вероятным.
Мы исключаем возможность карликового роста, вызванного неполноценным питанием, так как цыплята контрольной группы находились в тех же условиях вскармливания, света и т. д. и все же развивались совершенно нормально. Возможно, конечно, что внутризародышевое лечение вызывало изменение ассимиляции окружающей среды и, таким образом, даже в идентичных условиях цыплята не смогли закономерно метаболизировать полученные вещества. Это предположение, которое, думаем, заслуживает особого внимания, подтверждается также тем, что у одного из цыплят, подвергавшегося внутризародышевой обработке тироксином, наблюдался паралич, вызванный, вероятно, недостаточностью витамина В1, в то время как у соответствующего цыпленка контрольной группы, выращенного с ним в тех же условиях, не отмечалось подобных феноменов. М. Г. Закс в работе, посвященной роли щитовидной железы в эмбриональном развитии позвоночных, показывает, что значение тиреоидного гормона заключается в специальной регуляции метаболизма, а не в особом специфическом, как утверждает Нидем, формообразующем действии.
Во всяком случае, полученные нами результаты довольно интересны, так как мы не отмечали классического ускоряющего эффекта тироксина; при введении этого вещества наблюдалось не изменение длительности инкубационного периода, а как раз наоборот - парадоксальный эффект - появление карликового роста с незаращенными эпифизарными хрящами.
Как бы то ни было, экспериментальное воспроизведение нанизма подтверждает пластичность организмов и роль гормонов в эмбриональном развитии. Впрочем, многочисленные советские труды, появившиеся в результате научно-практической деятельности И. В. Мичурина, как например исследования, проведенные А. Д. Курбатовым, Г. А. Машталером и другими, предоставили нам богатый материал относительно возможности воздействия на развитие организма в различных стадиях внутризародышевой жизни.
Принимая во внимание полученные результаты посредством воздействия тироксина на зародыш развивающегося цыпленка, мы расширили наши исследования и на млекопитающих.
Клара Корд, которая много лет тому назад подвергала морских свинок в период беременности воздействию тироксина, отметила, что последние не рожают, и пришла к заключению, что зародыши резорбируются в матке. В действительности положение совсем иное.
Мы приступили к нашим ориентировочным опытам на трех группах морских свинок в состоянии беременности (всего на 23 морских свинках), которых подвергали ежедневно инъекциям тироксина. Это лечение мы проводили начиная с 5-го и по 15-й день беременности, причем вводили общую дозу в пределах 72-208 γ тироксина*.
* (Исследования были проведены совместно с М. Бэлэчану и Н. Албу.)
У всех подвергнутых воздействию тироксина морских свинок наблюдалось появление выкидыша. Выкидыш происходил в срок, обратно пропорциональный началу лечения. У морских свинок, подвергавшихся лечению с 5-го дня беременности, выкидыш происходил с большим запозданием, а именно всего за несколько дней до срока нормальных родов.
Морские свинки частично или полностью пожирали выкидышей, поэтому нам удалось произвести исследование лишь в одном случае. В этом наблюдении речь шла о морской свинке, у которой произошел выкидыш за 6 дней до срока, причем были абортированы 3 недоразвитые свинки с аномалиями головного мозга (как видно из рисунков) вплоть до анэнцефалии. Указанные плоды были примерно на 15 дней менее развиты, чем следовало бы по их хронологическому возрасту.
Указанные дефекты головного мозга, доходящие до экспериментальной анэнцефалии, выдвигают вопросы исключительной важности для изучения эмбрионального развития и факторов, могущих оказать влияние на последнее (рис. 202-207).
Особое значение невроэндокринных соотношений при эмбриональном развитии выявляется вследствие описанных нами фактов. Настоящими, а также ранее проведенными нами опытами, которые мы надеемся повторить и расширить, мы стремились показать, что гиперфункция щитовидной железы может играть определенную роль в появлении анэнцефалии.
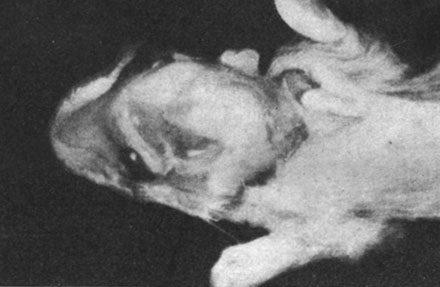
Рис. 204 Рис. 202, 203 и 204. - Выкидыши морских свинок за 6 дней до срока нормального рождения. Отмечается дефект мозгового вещества. В одном случае анэнцефалия
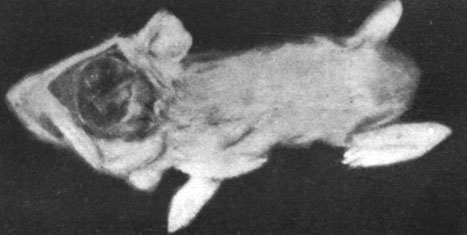
Рис. 204 Рис. 202, 203 и 204. - Выкидыши морских свинок за 6 дней до срока нормального рождения. Отмечается дефект мозгового вещества. В одном случае анэнцефалия
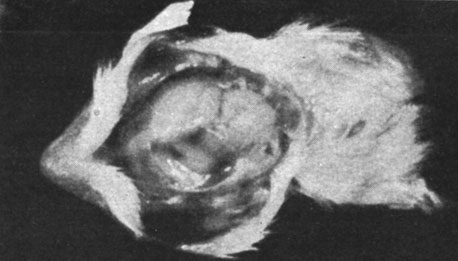
Рис. 204 Рис. 202, 203 и 204. - Выкидыши морских свинок за 6 дней до срока нормального рождения. Отмечается дефект мозгового вещества. В одном случае анэнцефалия
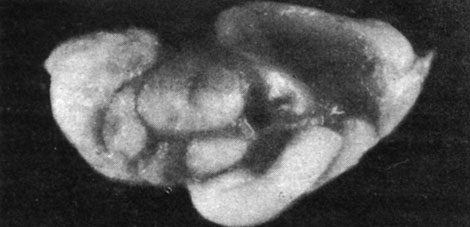
Рис. 205, 206 и 207. - Головной мозг животных, представленных на рисунках 202, 203 и 204
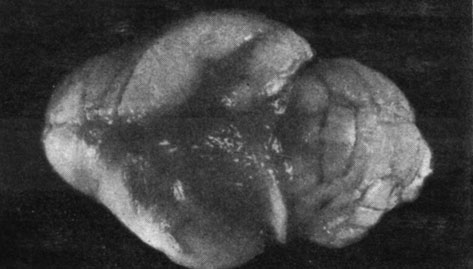
Рис. 205, 206 и 207. - Головной мозг животных, представленных на рисунках 202, 203 и 204
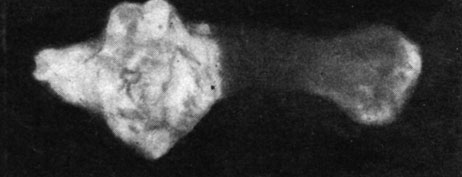
Рис. 205, 206 и 207. - Головной мозг животных, представленных на рисунках 202, 203 и 204
Нужно признать, что мы не ожидали получить столь интересных результатов. Изучая в связи с другой работой влияние базедовой болезни на потомство, мы настаивали на наличии наследственных уродств. У одних из представителей потомства наблюдались значительные соматические и психические расстройства, доходившие до имбецильности или идиотии.
В одном из случаев, находившихся под наблюдением Г. Маринеску, сын одной из больных базедовой болезнью был идиотом, а на секционном исследовании у него обнаружили весьма значительное утолщение черепной покрышки.
Однако нам ничего не известно из доступной нам медицинской литературы, что подтверждало бы наши настоящие наблюдения. С другой стороны, мы часто задаем себе вопрос, какие из эндокринных факторов воздействуют на развитие головного мозга. Было трудно допустить, что щитовидная железа может играть решающую роль, несмотря на то, что вышеприведенные факты, казалось бы, подтверждают столь исключительное значение щитовидной железы.
Было бы интересно исследовать средства, которые благоприятствуют, а не препятствуют развитию головного мозга. Возможно, что то же вещество, введенное в различных количествах, оказывает различное влияние; возникает вопрос, какие дозы тиреоидного вещества являются еще активно действующими и как оно действует на разные ткани и органы, и в частности, на кору головного мозга в различных фазах его развития: в детстве, у взрослого человека и у стариков различных возрастных периодов.
В том же направлении, т. е. в порядке изучения невроэндокринных соотношений в эмбриональном развитии, мы начали изучать воздействие тормозящих невротропных веществ нервной системы на развитие куриных зародышей. Мы применяли натриевую соль мединала (тормозящее нервную систему вещество и не оказывающее воздействия на рабочий орган, как показал еще Даниелополу) посредством ежедневных инъекций 1 γ в хориоаллантоисную оболочку, начиная с 6-го дня инкубационного периода. Наши исследования продолжаются; однако уже сейчас мы можем утверждать что у инъецированных цыплят наблюдался своевременный наклев и что у них не отмечалось никаких значительных морфологических или весовых изменений по отношению к контрольной группе.
При забитии одной из серий цыплят, через 2 дня после вылупления, мы получили интересные данные относительно веса некоторых органов.
Приводим ниже среднюю указанных органов подопытных цыплят в сопоставлении с контрольной группой:

Из вышеприведенных данных вытекает, что при внутризародышевой обработке натриевой солью мединала наблюдалась резкая гипотрофия нижнего мозгового придатка с различной гипотрофией других желез за исключением надпочечников, где наблюдалась гипертрофия. Точно так же следует отметить уменьшение веса печени и головного мозга.
Не будем обсуждать результатов. Оставим этот вопрос на более поздний срок, когда мы будем располагать более полными данными и указаниями относительно внезародышевого развития.
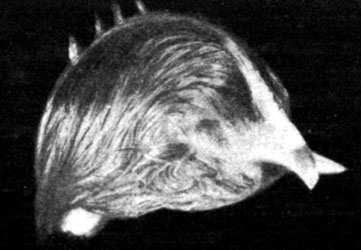
Рис. 208. - Вылупившийся в срок цыпленок с прижиганием глаза и перекрестом клюва
Эти исследования подтверждают значение невроэндокринных соотношений и вообще роль центральной нервной системы в гармоническом развитии организма.
Мы предприняли также с целью изучения роли нервной системы в эмбриональном развитии многочисленные исследования посредством метода прижиганий. Не так давно мы сообщили результаты, полученные посредством прижигания глаза, причем последний рассматривался как неврогенный орган. Благодаря вышеуказанному методу нам удалось получить атрофию лицевого костного массива со стороны поражения с перекрестом клюва и с уменьшением большого мозгового полушария с той же стороны (рис. 208). Впрочем, у спонтанных уродов, обнаруженных случайно, с отсутствием одного из глаз, наблюдались эти же аномалии.
В приведенных опытах отмечалось, что внутризародышевая обработка оказывает изменение на последующее развитие организма. Весьма знаменательным фактом является то, что, воздействуя на зародыш, можно изменить способ ассимиляции внешней среды во время незародышевой жизни, другими словами, видоизменяющий импульс не ограничивается первыми фазами развития, а продолжает влиять и впоследствии. Разумеется, при грубом воздействии применяемых факторов (прижиганий, разрушений и т. д.) эффект наблюдается непосредственно, однако нас интересует главным образом влияние, оказываемое на обмен веществ.
Богатый советский опыт, основанный на учении И. В. Мичурина, а также несколько фактов, отмеченных нами в том же направлении, дают нам право считать, что средняя длительность жизни какого-либо рода животных, имеющая в настоящий момент статистически достоверное значение, может быть надолго изменена посредством воздействия на ряд поколений во время зародышевого развития.
|
ПОИСК:
|
При использовании материалов проекта обязательна установка активной ссылки:
http://gelib.ru/ 'Геронтология и гериатрия'