1. Рыбы
"Недетерминированный" рост у рыб, который послужил Биддеру основой для его гипотезы, часто служил предметом всестороннего обсуждения [501, 550, 596, 1187, 1222, 1255]. Многие крупные виды костистых рыб могут расти на протяжении всей жизни, а скорость роста у них снижается значительно менее резко, чем, например, у большинства рептилий. Классическим примером непрерывного роста без признаков старения, оцениваемого статистически или по способности к размножению, служат самки камбал. В данном случае факты свидетельствуют в пользу гипотезы Биддера, так как у самцов рост прекращается сравнительно рано и имеются данные, что продолжительность жизни у них короче, чем у самок [1281]. В то же время у многих мелких костистых рыб установлено старение, выражающееся в снижении способности к размножению, и хотя самки могут расти всю жизнь, видовой возраст существует, по-видимому, у обоих полов. Аквариумисты знают, что многие костистые рыбы с возрастом теряют способность к размножению. У отдельных видов, по-видимому, имеется предельный возраст, но до сих пор не опубликовано ни одной таблицы выживания рыб в аквариуме, которая могла бы подкрепить это впечатление. Изучен рост только некоторых мелких костистых рыб [341, 1255] (фиг. 20).
![Фиг. 20. Рост рыб (в длину) в течение одного года жизни [1255]. А. Самцы. Б. Самки. I - Xiphophorus; II - Lebistes; III - Heterandria. Знак или означает наступление половой зрелости](pic/000031.jpg)
Фиг. 20. Рост рыб (в длину) в течение одного года жизни [1255]. А. Самцы. Б. Самки. I - Xiphophorus; II - Lebistes; III - Heterandria. Знак ♂ или ♀ означает наступление половой зрелости
Судя но наблюдениям за природными популяциями, мелкие костистые рыбы, а может быть, и все костистые рыбы вообще проявляют старение, оцениваемое как статистически, так и по снижению способности к размножению, причем старение этих рыб напоминает старение млекопитающих [398, 399]. Наиболее догматические утверждения в этой области принадлежат патологоанатомам. Обнаружение в семенниках одного из видов костистых рыб (Astyanax americcmus) конкрементов Расквин и Хафтер [963] рассматривают как "появление старческих изменений, свидетельствующее о том, что вопреки широко распространенной неправильной концепции костистые рыбы подвержены старению общего типа, характерному для всех позвоночных". Снижение плодовитости у некоторых аквариумных видов рыб служит более основательным подтверждением такой точки зрения. Во всяком случае, у многих видов рыб наблюдаются своеобразные колебания величины смертности, связанной с размножением. Трактовать эти колебания величины смертности как особую форму старения у тех видов, которые после размноженин всегда гибнут (например, самцы Callionymus [188] и миноги), затруднительно, поскольку у других рыб после каждого сезона размножения некоторое ограниченное число особей выживает и после этого получает еще на какой-то срок "право на жизнь". Сомнительно, чтобы какие-либо циклические или потенциально циклические изменения смертности можно было назвать старческими. У Callionymus lyra в естественных условиях самец, по-видимому, живет до 6 лет, а самка - до 7 лет. Самцы после единственного сезона размножения исчезают; вероятно, они гибнут, а возможно, мигрируют на большую глубину. Самки впервые нерестятся на 3-, 4- или 5-м году жизни в зависимости от скорости роста и, вероятно, размножаются в течение ряда лет [188]. В таком случае у запаздывающих в своем развитии самок общая продолжительность жизни, вероятно, будет больше.
![Фиг. 21. Рост форели в оз. Уиндермир и в небольших горных озерах [383]. I-III - рост в длину; I - в оз. Уиндермир; II - в оз. Три-Даб-Тарн; III - в оз. Ю-Три-Тарн; IV - увеличение веса; в оз. Три-Даб-Тарн](pic/000032.jpg)
Фиг. 21. Рост форели в оз. Уиндермир и в небольших горных озерах [383]. I-III - рост в длину; I - в оз. Уиндермир; II - в оз. Три-Даб-Тарн; III - в оз. Ю-Три-Тарн; IV - увеличение веса; в оз. Три-Даб-Тарн
Природные популяции почти всегда изучают в условиях, когда сила смертности, устанавливаемая для всей жизни, очень высока, и поэтому такие исследования дают мало сведений об изменениях смертности в поздние периоды жизненного цикла долгоживущих форм. Если исключить очень высокую смертность личиночных форм, то популяции многих видов рыб, изучаемых в природных условиях, по возрастному составу и характеру смертности подобны птицам, т. е. высокая и постоянная смертность не зависит от возраста, а продолжительность предстоящей жизни фактически не меняется [284, 383]. Существенные различия в продолжительности жизни, возможно, обусловлены наличием пригодных кормовых организмов, подходящих по своим размерам для питания взрослых, и конкуренцией между молодью изучаемых видов и взрослыми формами других. У некоторых популяций Pimephales видовой возраст явно во много раз ниже, чем в популяциях близкородственных видов [380, 11661. По данным Маркуса [765], у черноголовой пимефалес (Pimephales promelas) все особи достигали видового возраста и видовых размеров; исключение составляли немногие особенно крупные индивидуумы. Очевидно, это обусловлено тем, что после нереста гибнет 80% особей; таким образом, выжившие особи и особи, не принимавшие участия в нересте в первый год зрелости, продолжали расти до следующего сезона размножения. Если размножения не произошло, жизнь особи продолжается. Биддер [97] отмечает, что угри, которые, как полагают, обычно гибнут после нереста, в неволе живут долгие годы [353]. Фрост и Смайли [383] обнаружили значительные различия в скорости роста и в форме кривой роста у форелей, обитающих в небольших горных озерах и в озере Уиндермир (фиг. 21). Возрастной состав популяции форели в горных озерах вполне соответствует постоянному уровню выживания 35% в год между 2- и 8-м годами жизни. В этих озерах рост рыб сильно замедлялся, в озере же Уиндермир отдельные особи еще росли до 7 лет приблизительно по линейному закону. Способность к дальнейшему росту может быть связана с достижением размеров, позволяющих охотиться на более мелких рыб. Долгоживущие рыбы, такие, как щука, несомненно, растут очень долго [1045], но увеличение размеров их тела связано с увеличением размеров их добычи [381]. Риккер [979] замечает, что "смерть от старости - банальное событие" в популяциях ушастого окуня. Этот вывод, однако, был вызван тем, что не удалось обнаружить гибели рыб в результате действия обычных факторов (болезни и хищников). Общий уровень смертности, установленный в экспериментах с введением метки, составил 56% для мелких особей и 58% для более крупных (более старых). Во многих популяциях других видов, не являющихся промысловыми, с увеличением возраста и размеров смертность постепенно повышается [980]. Геркинг [398] собрал веские доказательства старения во многих природных популяциях рыб и дал сводку литературы. Особо интересный случай - гибель родителей у лососевых, которые умирают после нереста от эндокринных изменений, сопутствующих миграции. Немигрирующие популяции выживают после нереста, и входящие в их состав особи размножаются многократно. Робертсон тщательно изучил этот вопрос и в своих сводках приводит относящуюся к нему литературу [986-988]. Предполагаемая причина родительской смерти - гиперфункция надпочечников; однако наблюдаемые изменения очень похожи на происходящие в старости у тех рыб, гибель которых после миграции предотвращали путем кастрации в раннем возрасте.
![Фиг. 22. Константы роста тела в длину для севрюги (Acipenser stellatus) в зависимости от возраста [1048]. Ломаная линия - данные для самок; отдельные кружки - данные для самцов; сплошная линия - усредненные данные для самцов; пунктирная линия - усредненные данные для самок. K1 = 0,67; К2=0,58](pic/000033.jpg)
Фиг. 22. Константы роста тела в длину для севрюги (Acipenser stellatus) в зависимости от возраста [1048]. Ломаная линия - данные для самок; отдельные кружки - данные для самцов; сплошная линия - усредненные данные для самцов; пунктирная линия - усредненные данные для самок. K1= 0,67; К2=0,58
Шмальгаузен [1018] собрал много важных сведений из работ ряда русских авторов (например, [1183]). У севрюги активный рост продолжается по крайней мере в течение 30 лет с небольшим падением скорости в период полового созревания (около 16 лет). У леща скорость роста уменьшается более равномерно. Эти рыбы достигают половой зрелости примерно к 3 годам, а дегенеративные изменения в половых железах обнаруживаются у них обычно начиная с 6-го года; на кривой роста [101] можно выделить две четкие стадии: одна начинается после достижения половой зрелости, а другая - после возникновения старческих изменений в половых железах. Константа роста снижается до некоторой величины, после чего не меняется до 13-летнего возраста (фиг. 22-24). Такой тип роста довольно близко совпадает с типом роста, описанным для золотой рыбки.
![Фиг. 23. Возрастные изменения веса тела у севрюги {Acipenser stellatus) (I) и леща (Abramis brama) {II) [1048]. Одно деление шкалы веса для севрюги равно 4 г, а для леща - 1 г.](pic/000034.jpg)
Фиг. 23. Возрастные изменения веса тела у севрюги {Acipenser stellatus) (I) и леща (Abramis brama) {II) [1048]. Одно деление шкалы веса для севрюги равно 4 г, а для леща - 1 г.
Интересные кривые выживания для ряда видов рыб приводят Бивертон и Холт [95] (фиг. 25).
У Xiphophorus и Lebistes самцы характеризуются строго определенными видовыми размерами, но самки продолжают заметно расти на протяжении всей жизни, так что по типу роста они мало отличаются от камбал [1255]. Однако более ранние опыты, проведенные на этих рыбах, указывают на отсутствие резких различий в выживаемости самцов и самок при разведении в аквариуме [78]. У Heterandria и те и другие фактически достигают предельных размеров [1255] (фиг. 20, А и Б). У золотой рыбки при разведении ее в аквариуме плодовитость достигает максимума на 3-й год жизни, после чего снижается, и к 7-му году почти все особи оказываются бесплодными. Отмечен, правда, случай размножения в 10- летнем возрасте [511]. Однако по окончании репродуктивного периода состояние рыбы иногда значительно улучшается и она оказывается значительно менее чувствительной к неблагоприятным внешним воздействиям. У декоративных рыб продолжительность жизни равна примерно 17 годам, хотя известны и гораздо более старые экземпляры. Максимальная продолжительность жизни составляет 30-40 лет. Скорость роста крайне изменчива. Одна особь, содержавшаяся 25 лет в 20-литровом резервуаре, достигла в длину только 10 см [511].
![Фиг. 24. Возрастные изменения веса тела у леща (Abramis brama) [1048]. Средние константы роста на различных стадиях жизни (неполовозрелый, репродуктивный и пострепродуктивный периоды) показаны горизонтальными линиями К1, К2 и К3](pic/000035.jpg)
Фиг. 24. Возрастные изменения веса тела у леща (Abramis brama) [1048]. Средние константы роста на различных стадиях жизни (неполовозрелый, репродуктивный и пострепродуктивный периоды) показаны горизонтальными линиями К1, К2 и К3
Ни в одном из приведенных случаев не было выяснено, какая часть потенциального жизненного цикла действительно отражена в наблюдаемой кривой роста. У большинства рыб скорость роста фактически уменьшается с возрастом, хотя у многих из них эффективный репродуктивный период прекращается задолго до того, как это уменьшение приведет к почти стабильным размерам тела. Более того, падение способности к размножению, очевидно, не подразумевает ослабления жизненности и в действительности может вызвать противоположный эффект, поскольку размножение многих рыб связано с целым рядом опасностей.
![Фиг. 25. Примеры кривых выживания для относительно маловылавливаемых популяций рыб [95]. А. Сельдь (Clupea); I - С. sprattus; II - С. pallasi; III - С. harengus. Б. Cottus gobio. В. Окунь Perea fluvatilis. Г. Callionymus lyra. Д. I - Leuresthes tenuis; II - Leucichthys kiyl; III - Coregonus clupeaformis; IV - Aeipenser fulvenscens. N число выживших](pic/000036.jpg)
Фиг. 25. Примеры кривых выживания для относительно маловылавливаемых популяций рыб [95]. А. Сельдь (Clupea); I - С. sprattus; II - С. pallasi; III - С. harengus. Б. Cottus gobio. В. Окунь Perea fluvatilis. Г. Callionymus lyra. Д. I - Leuresthes tenuis; II - Leucichthys kiyl; III - Coregonus clupeaformis; IV - Aeipenser fulvenscens. N число выживших
Ответ на большинство этих вопросов, очевидно, можно получить с помощью прямых экспериментов на мелких видах костистых рыб. Для этих целей особенно подходят гуппи (Lebistes), так как их легко разводить и измерять, а также регулировать их рост, варьируя количество корма и жизненное пространство.
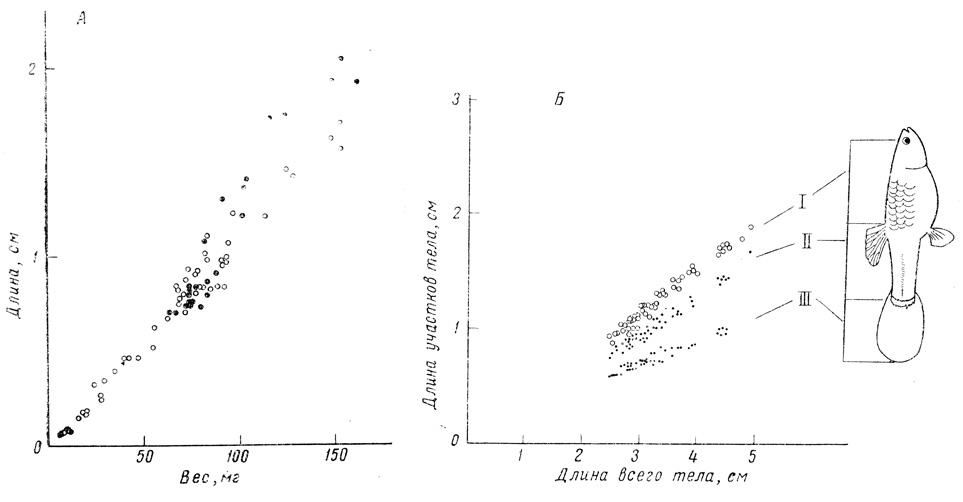
Фиг. 26. Рост самок групп (Lebistes). А. Соотношение веса и длины тела; ○ - непрерывный рост, ● - задержанный и возобновленный рост; разброс в старших возрастах обусловлен сохранением яиц у некоторых рыб. Б. Изометрия роста при задержке. По абсциссе отложена длина всего тела, а по ординате - длина одного из трех участков: I - от середины глаза до переднего края спинного плавника; II - от переднего края спинного плавника до корня хвоста; III - от корня хвоста до его конца
Ограничивая жизненное пространство и диету, можно затормозить рост самок гуппи примерно на протяжении 600 дней, так что их длина будет составлять всего лишь около 2 см. В таком состоянии они сохраняют способность размножаться (в отличие от крыс с заторможенным ростом в опытах Мак-Кея; см. стр. 230) и возобновлять свой рост. Для каждого резервуара и уровня питания характерны свои "видовые размеры"; другими словами, при каждой плотности популяции при переносе рыб в более просторный аквариум или при удалении части рыб на кривой роста быстро достигается новое плато. На кривых, приведенных Велленсиком, отражен только один пример такой стабилизации. У гуппи способность к росту с возрастом, по-видимому, также несколько снижается, и размеры тела практически имеют предел, достигаемый при максимальном жизненном пространстве и максимальном снабжении пищей.
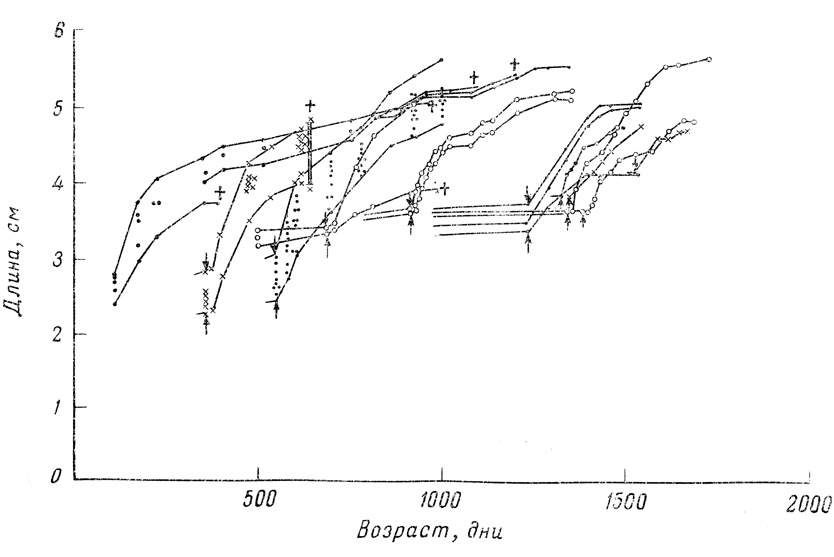
Фиг. 27. Рост самок гуппи с задержкой. Полноценное кормление возобновляли в момент, указанный стрелкой, а смерть показана крестиком
Тот факт, что, согласно последним данным, продолжительность жизни у нерастущих самцов и растущих самок значительно не различается, позволяет предполагать, что большое количество сведений о росте и старении у рыб можно получить путем сбора статистических данных о гуппи с различной скоростью роста.
В 1950 г. было предпринято популяционное изучение гуппи (Lebistes reticulatus Peters) для получения ответа па вопрос, увеличивается ли с возрастом смертность этих рыб в различных условиях их содержания и как это увеличение связано с ростом [241, 244, 245, 247] (фиг. 26- 33).
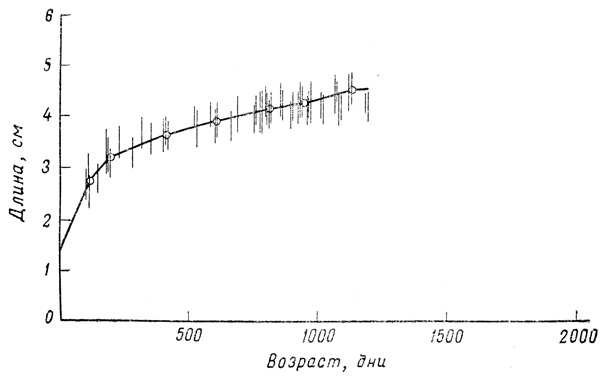
Фиг. 28. Рост самок гуппи в 'нормальных' аквариумных условиях, обеспечивающих медленный непрерывный рост. Вертикальные линии показывают пределы для отдельных аквариумов, а кривая отражает средние величины для всей популяции (48 особей)
Основной результат статистического исследования можно выразить довольно четко: наблюдаемые кривые выживания гуппи как по своей форме, так и по изменчивости под влиянием условий среды не сильно отличаются от подобных же кривых, полученных для мелких млекопитающих в лабораторных условиях. Во всех сериях и при всех экспериментальных условиях сила смертности неуклонно увеличивается с возрастом. При улучшении условий содержания кривая выживания все больше и больше приобретает прямоугольную форму, и варианса уменьшается, но в конце кривой образуется своего рода выступ, так что создается типичный "параллелограмм выживания", характерный для животных с определенной продолжительностью жизни (фиг. 32).
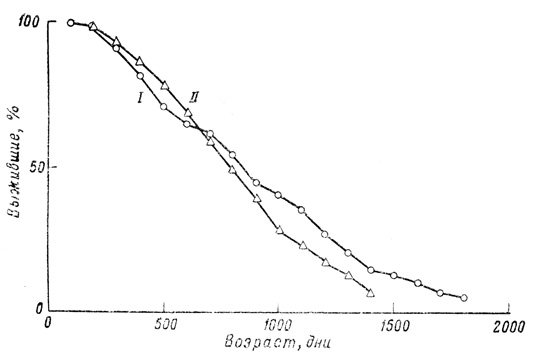
Фиг. 29. Обобщенные кривые выживания для самок (351 особь) (!) и самцов (312 особей) (II) из всех аквариумов
Старение у гуппи, очевидно, проявляется при сохранении способности к росту. Это противоречит гипотезе Биддера [97]; при отсутствии данных о постоянном обновлении всех клеток нельзя отвергнуть теории, основывающиеся на отмирании клеток; например, гистологическая картина дегенеративных изменений в почках не зависит от того, продолжается рост или нет. Остается спорным, оказывают ли благоприятные для роста условия такое же влияние на старение у долгоживущих рыб, например у севрюги, как и у гуппи. Самки гуппи при благоприятных условиях роста достигают 75% своих предельных размеров (L ", по терминологии Бивертона и Холта [95]) в возрасте 700 дней, а их предельный возраст (Хос ), по-видимому, составляет примерно 2000 дней; наши самые старые рыбы доживали до 2200 дней. Соответствующей величины L" камбала достигает в 14 лет ([95], стр. 284), что при сохранении того же соотношения дает максимальную продолжительность жизни 40 лет. На кривой роста севрюги не обнаруживается ни малейшего уплощения в точке, соответствующей 30 годам [1048]. Допустим, что севрюга к 45 годам вырастает до 75% максимальной длины, достигаемой асимптотически; исходя из этих цифр, можно ожидать, что величина приближается к 128 годам. Для севрюги рекордный возраст не менее 82 лет был установлен по срезам плавника.
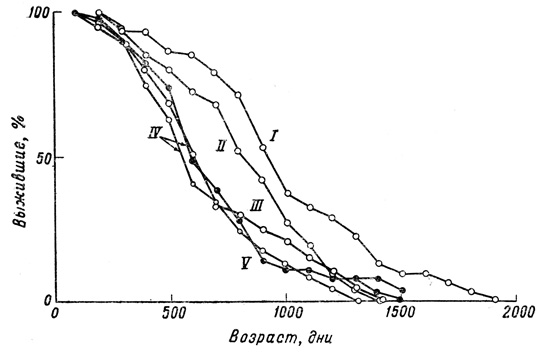
Фиг. 30. Кривые выживания для самцов гуппи, содержавшихся в больших аквариумах (I-III), 600-миллилитровых банках (IV) и 250-миллилитровых бутылках из-под молока (F)
Используя данные для гуппи, можно и для других видов рыб построить кривые, иллюстрирующие изменение жизнеспособности (при тех же соотношениях между ростом и выживанием). На фиг. 34 приведены кривые роста для 87 самок (содержавшихся по партиям), выведенных в аквариуме, вместе с кривой выживания для всей популяции на протяжении 1100 дней.
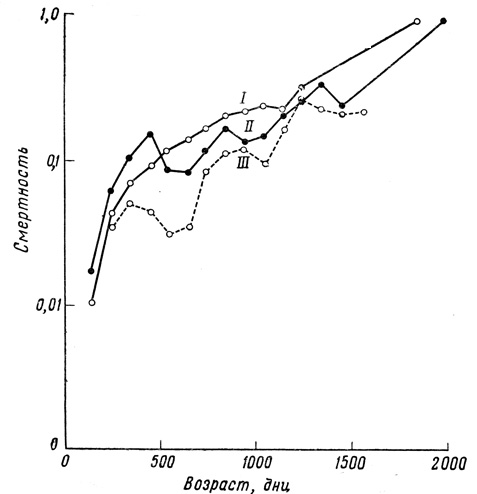
Фиг. 31. Изменение смертности гуппи с возрастом в больших аквариумах. I - самцы; II - самки; III - отдельные случаи гибели от эпидемии
На графиках, приведенных на фиг. 35, рост четырех видов рыб (колюшка, треска, камбала, хек) в природных условиях отложен в зависимости от времени в пределах кривой для гуппи (поправочные коэффициенты перевода шкалы времени: для колюшки 0,91; для трески 3,65; для хека 7,3; для камбалы 10,95). Если эти соотношения сохраняют силу, то следует ожидать, что популяции с такими кривыми роста будут уменьшаться на четверть только по причине старения соответственно к 2,5, 10, 20 и 30 годам, а предельные возрасты для этих четырех видов рыб составят 5, 20, 40 и 60 лет. Колюшка в естественных условиях фактически живет 2-3 года, и лишь немногие особи живут больше. У других видов промысловых рыб, если бы старение было связано с ростом, как У гуппи, смертность не была бы значительной в пределах обычной ожидаемой продолжительности жизни; модальный возраст камбалы в уловах составляет 6 лет. Величины предельного возраста выглядят также приемлемыми, за исключением того, что если исходить из одних только размеров тела, то продолжительность жизни трески должна быть выше 20 лет.
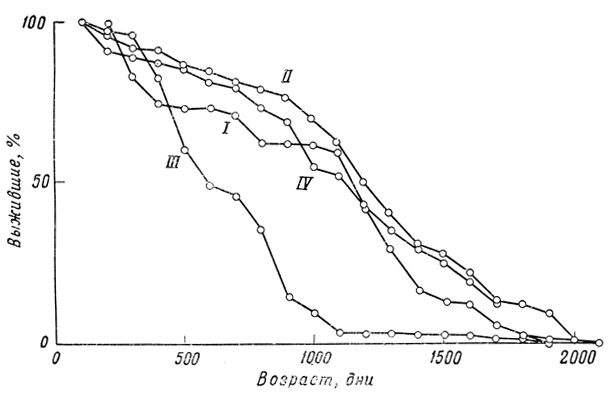
Фиг. 32. Кривые выживания для самок гуппи, содержавшихся в больших аквариумах (I-III) и для самок, отсаженных в 2-литровые банки (IV)
Таким образом, исходя из существующих данных, нет никаких оснований предполагать различные типы смертности для мелких и крупных рыб.
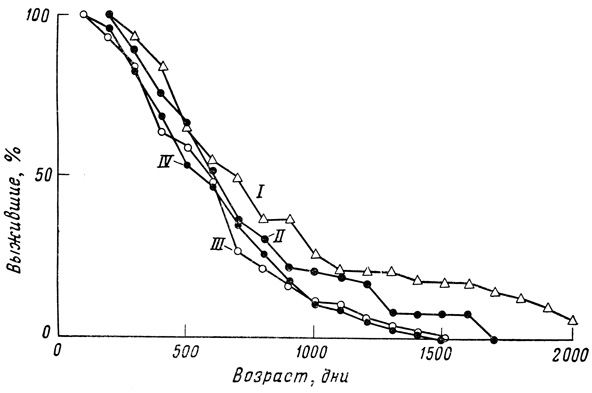
Фиг. 33. Кривые выживания для самок гуппи, содержавшихся в 2-литровых банках (I), 600-миллилитровых банках с полноценным питанием (II), в 600-миллилитровых банках с ограниченным питанием (III) и в 250-миллилитровых бутылках из-под молока (IV)
Представляется вполне вероятным, что у костистых рыб имеется столько же разновидностей "старения", сколько типов роста. Некоторые формы, очевидно, напоминают монокарпические растения; смертность у них связана с размножением. У других (при содержании в аквариумах) продолжительность жизни определяется старением, а их смертность с возрастом увеличивается по кривой, весьма напоминающей соответствующую кривую для млекопитающих. Можно допустить, что у ряда форм продолжительность жизни действительно не детерминирована, однако это будет означать лишь, что "детерминированный" предельный срок для них, как и для птиц в природных условиях, наступает слишком поздно и практически никогда не достигается.
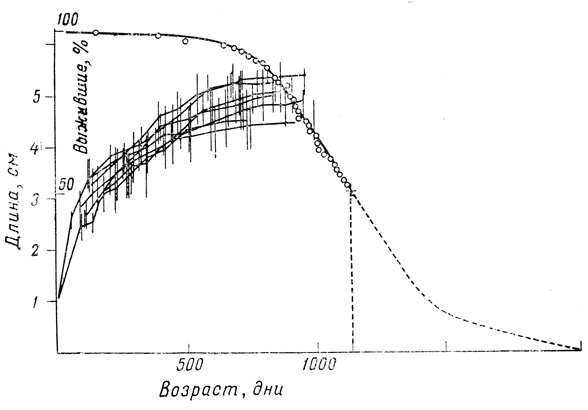
Фиг. 34. Рост и выживание самок гуппи. Ср. с фиг. 35
|
ПОИСК:
|
При использовании материалов проекта обязательна установка активной ссылки:
http://gelib.ru/ 'Геронтология и гериатрия'